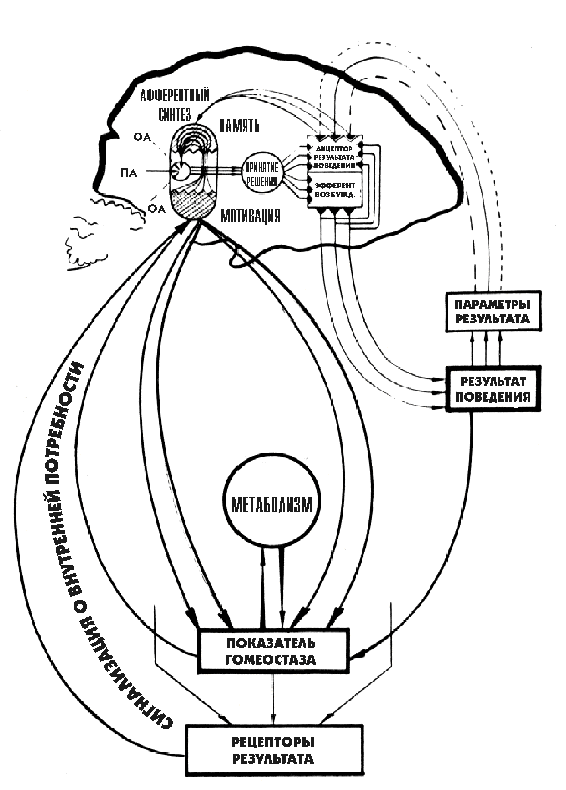
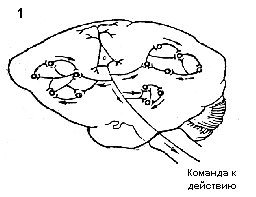
Рис 1. Общая схема функциональной системы по П.К.Анохину,
УДК 519.7
На 26-м Международном конгрессе физиологических наук (ICPS) в Нью-Дели (Индия) в 1974 году известный американский нейропсихолог Самуил Корсон в пленарном докладе, посвященном памяти П.К.Анохина, публично заявил, что П.К.Анохин по праву признан основоположником физиологической кибернетики. Норберт Винер во время своего визита в Москву в 1966 году и посещения Сеченовско-го института физиологии, которым руководил П.К. Анохин, также признал, что работы П.К. Анохина по физиологической кибернетике намного опередили зарождение кибернетического направления в других отраслях науки.
Как известно, Н.Винер, которого считают Lотцом кибернетикиҰ, в 1948 году теоретически определил кибернетику как Lуправление и связь в животном и машинеҰ [8, 16].
П.К.Анохин в 1932-35 годах, будучи заведующим кафедрой физиологии сначала в Нижегородском Университете, а затем - в Горьковском медицинском институте, совместно с коллективом своих молодых сотрудников и студентов активно экспериментально исследовал проблему соотношения центров и периферии в нервной деятельности с помощью оригинальной, разработанной им методики гетерогенных анастомозов нервов [1]. Метод гетерогенных анастомозов заключался в том, что у животных осуществлялась операция перерезки нервных стволов, а затем центральный конец какого-либо предварительно перерезанного нерва подшивался к периферическому концу чужеродного ему нерва. Вследствие такой операции нейрофибриллы центрального отрезка нерва прорастали в ствол чужеродного нерва и на периферии афферентные волокна устанавливали новые контакты с неадекватными для них рецепторами, а эфферентные волокна устанавливали функциональный контакт с неадекватной для соответствующего нервного центра мышечной, железистой или нервной тканью. В результате создавались своеобразные экспериментальные LхимерыҰ, у которых при раздражении периферических рецепторов можно было вызвать неадекватные для связанного с ними прежде нервного центра реакции и в то же время наблюдать у животных неадекватные прежнему нервному центру эффекторные реакции. Примером гетерогенного анастомоза, широко применявшегося в лаборатории П.К.Анохина, является анастомоз центрального конца перерезанного на шее правого блуждающего нерва с периферическим отрезком перерезанного смешанного нерва передней конечности собаки, например срединного или лучевого нерва. В других случаях центральный конец блуждающего нерва подшивали к периферическому отрезку перерезанного, в основном, эфферентного нерва барабанной струны (n. сhorda tympany), ранее иннервировавшего подчелюстную слюнную железу, или к периферическому концу, в основном, афферентного язычного нерва (n. lingualis). По прошествии 3-х - 4-х месяцев после таких операций нейрофибриллы центрального отрезка блуждающего нерва активно прорастали в стволы искусственно связанных с ними через анастомозы периферических отростков и устанавливали функциональный контакт в первом случае - афферентные волокна блуждающего нерва - с рецепторами кожи, а эфферентные его волокна, включая входящий в ствол блуждающего нерва возвратный нерв (n. reccurrens), - c проприоцепторами соответствующих мыщц передней конечности собаки. При второй операции эффекторные волокна блуждающего нерва прорастали в ствол нерва барабанной струны и устанавливали функциональный контакт с подчелюстной слюнной железой. При третьей операции афферентные волокна блуждающего нерва прорастали и устанавливали функциональный контакт со вкусовыми рецепторами языка. В первом случае при легком почесывании участка кожи, иннервируемого искусственно благодаря анастомозу блуждающим нервом, у собак возникал кашель. При прикладывании к этому участку слегка десквамированной кожи растворов хлористого натрия у животных возникали сокращения желудка и рвота. Во втором случае LвагуснаяҰ слюнная железа изменяла свою секрецию в соответствии с процессом пищеварения. В третьем случае блуждающий нерв восстанавливал свойства рецепторов сосочков языка, и при их химическом раздражении тоже можно было влиять на ядро блуждающего нерва.
Указанные необычные явления при целостности второго блуждающего нерва сохранялись без изменений долгое время. Это отличало анастомозы блуждающего нерва от гетерогенных анастомозов смешанных нервов, иннервирующих мышцы с противоположными функциями, например агонисты и антагонисты. При этих анастомозах в течение нескольких месяцев у животных восстанавливались нарушенные функции ходьбы.
Эти и другие эксперименты заставили П.К.Анохина обратить особое внимание на решающую роль поступающей с периферии афферентации в перестройке функций нервных центров.
На основании этих экспериментов П.К.Анохин уже в начале 30-х годов нашего столетия формулирует понятие Lфункциональная системаҰ. Функциональные системы по П.К.Анохину - динамические саморегулирующиеся организации, все составные компоненты которых взаимосодействуют достижению полезных для функциональной системы и организма в целом приспособительных результатов. В качестве элементов, строящих функциональные системы, выступают различные показатели внутренней среды организма, поведения и психической деятельности человека, удовлетворяющие его исходные потребности.
В функциональных системах П.К.Анохин отчетливо подчеркнул ведущую LтворческуюҰ роль афферентации, поступающей от периферических органов - результата действия - в соответствующие нервные центры. Эту афферентацию П.К.Анохин за 12 лет до Н.Винера назвал Lобратной афферентациейҰ.
В книге LБиология и нейрофизиология условного рефлексаҰ [5] П.К.Анохин писал: LОдним из важнейших условий целостности функциональной системы как интегративного образования организма, включающего в себя центральные и периферические образования, мы считаем наличие обратной афферентации о достигнутом конечном приспособительном эффекте. Это позволило нам рассматривать функциональную систему как замкнутое физиологическое образование с непрерывной обратной информацией об успешности данного приспособительного действия [1]. В сущности, в этой теории были предвосхищены все основные черты кибернетического замкнутого контура с обратной связью, т. е. Lfeed backҰ, как она была названа позднееҰ.
Роль обратной афферентации отчетливо выступила и в других опытах, выполненных П.К.Ано-хиным, в частности, с подменой подкрепления в условнорефлекторных экспериментах [2].
В этом же направлении П.К.Анохина заставляли думать и эксперименты с компенсацией движений у животных в случае удаления одной или нескольких конечностей. Отсутствие конечностей приводило в этих случаях к реорганизации характера движения животных, постоянно стимулировало перестройку их двигательных функций и создавало новые условия к движению с помощью оставшихся конечностей.
Значение афферентации, идущей от соответствующей периферии, в деятельности нервных центров было убедительно продемонстрировано П.К.Анохиным и его сотрудниками и в экспериментах с пересадками конечностей в эмбриогенезе у аксолотлей [3].
Таким образом, можно видеть, что П.К.Анохин, в отличие от Н.Винера, сформулировал кибернетические закономерности на основе многочисленных экспериментов. П.К.Анохин выявил роль Lобратной афферентацииҰ экспериментально как неотъемлемое, объективно существующее свойство открытых им функциональных систем организма.
Парадоксально, что как П.К.Анохин, так и позднее Н.Винер пришли к формулированию кибернетических закономерностей только в середине нашего столетия, в то время как живая природа LнащупалаҰ эти закономерности многие миллионы лет тому назад и использовала их в построении различных функциональных систем организмов.
Функциональные системы, открытые П.К.Ано-хиным, являются объективно существующей реальностью. Они воплощают в своей деятельности ведущие кибернетические свойства обратной связи и регулирования по заданному параметру. В функциональных системах обратная афферентация (она же обратная связь) включена в звенья оценки функциональными системами исходных потребностей и их удовлетворения путем достижения полезных для систем и организма в целом результатов.
П.К.Анохиным разработаны новые представления о центральной архитектонике системной организации поведенческих актов, включающей стадии афферентного синтеза, принятия решения, предвидения потребного результата (акцептор результата действия), эфферентный синтез и целенаправленное действие, ведущее к потребному результату. Параметры достигнутых результатов на основе обратной афферентации оцениваются по П.К.Анохину акцептором результата действия (рис.1).
Теорией функциональных систем П.К.Анохин постулировал новый принцип жизнедеятельности - принцип системной организации физиологических функций, который наряду с рефлекторным принципом прочно занял свое место в физиологии [10].
В последние годы теория функциональных систем получила дальнейшее развитие в концепции системного квантования процессов жизнедеятельности [11, 12]. В соответствии с концепцией системного квантования континуум различных процессов жизнедеятельности подразделяется постоянно формирующимися потребностями живых организмов и их удовлетворением на специальные Lсистемо-квантыҰ. Каждый LсистемоквантҰ строится соответствующей функциональной системой. LСистемо-квантыҰ проявляются на различных уровнях жизнедеятельности: метаболическом, клеточном, в процессах гомеостазиса и поведения.
LСистемоквантыҰ инициируются исходной потребностью,
и в случае поведенческой деятельности организма, - соответствующей
мотивацией. Наряду с этим в них включается деятельность, направленная
на удовлетворение исходной потребности, и постоянная оценка промежуточных
и конечного результатов деятельности с помощью обратной афферентации.
Нетрудно заметить, что в LсистемоквантахҰ обратная афферентация
также включается в звено оценки потребности и в звено оценки параметров
промежуточных и конечного результатов, удовлетворяющих эту потребность
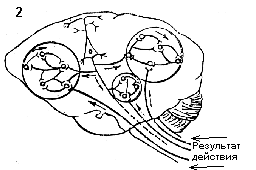
(рис. 2).
Рис 1. Общая схема функциональной системы по П.К.Анохину,

Рис.2. Обратная афферентация
в LсистемоквантахҰ, определяющих оптимальный для метаболизма показатель
гомеостазиса (Р) за счет внутреннего и внешнего (поведенческого)
звеньев саморегуляции. Р1 - отклонение результата от
уровня, определяющего нормальный метаболизм (М). РВ1
и РВ2 - промежуточные результаты вегетативной деятельности,
направленной на нормализацию показателя гомеостазиса. РП1,
РП2, РП3 - промежуточные результаты поведенческой
деятельности, ведущие к восстановлению оптимального уровня показателя
гомеостазиса. Обратная афферентация показана утолщенными стрелками.
П.К.Анохин, в отличие от других специалистов в области кибернетики, обнаружил в функциональных системах аппарат оценки потребности и ее удовлетворения, который был назван им Lакцептором результата действияҰ.
Акцептор результата действия представлен в функциональных системах различного уровня организации, начиная от молекулярных до сложноорганизованных психических процессов. Акцептор результата действия сложился в определенную мозговую архитектонику в функциональных системах поведенческого, а у человека - психического уровня. Акцептор результата действия на информационной основе опережающе заключает в себе генетически детерминированные и индивидуально приобретенные свойства параметров результатов деятельности, удовлетворяющих исходные потребности организма.
На всех уровнях организации акцептор результата действия представлен молекулярными энграммами, постоянно совершенствующимися в процессе индивидуальной деятельности живых существ.
Наши исследования показали, что, под влиянием подкрепления, т. е. удовлетворения исходной потребности, нейроны, вовлеченные в доминирующую мотивацию, начинают активно экспрессировать информационные молекулы, образующие своеобразную LэнграммуҰ подкрепления [13].
Обратная афференция и акцептор результата действия в каждой функциональной системе представляют таким образом важнейшие звенья их саморегуляторной деятельности. Благодаря акцептору результата действия в каждой функциональной системе посредством обратной афферентации постоянно осуществляется оценка состояния ее полезного для организма результата и происходит коррекция деятельности по удовлетворению исходных потребностей в разных условиях существования субъектов.
Функциональные системы, будучи продуктом длительного эволюционного развития живых организмов, воплотили в своей деятельности все известные в настоящее время кибернетические закономерности.
Наиболее важным кибернетическим свойством функциональных систем является их самоорганизация. В последние годы процессы самоорганизации стали рассматривать в качестве отдельного раздела науки - синергетики.
Самоорганизация присуща каждой составляющей функциональной системе организма.
Самоорганизация функциональных систем начинается с эмбрионального, пренатального и раннего постнатального периодов развития плода.
Процессы самоорганизации функциональных систем составляют специальный раздел общей теории функциональных систем, названный П.К.Ано-хиным LсистемогенезомҰ. Под системогенезом П.К.Анохин предложил понимать процессы становления, развития и усовершенствования функциональных систем и их отдельных частей в пре- и постнатальном онтогенезе.
Уже в пренатальном онтогенезе в соответствующих тканях плода на генетически детерминированной основе опережающе созревают специальные рецепторы. На эти рецепторы начинают действовать информационные молекулы - гормоны, олигопептиды и другие биологически активные вещества, циркулирующие в крови матери. Под влиянием этих молекул в утробе материнского организма у плода складываются функциональные системы гомеостатического уровня.
Другая группа рецепторов опережающе созревает в онтогенезе по отношению к полезным приспособительным результатам поведения, которые на ранних этапах постнатального развития играют решающую роль для выживания новорожденных животных. В этом случае на основе механизма запечатления (импринтинга) происходит формирование функциональных систем поведенческого уровня.
Изложенное свидетельствует о том, что полезные приспособительные результаты выступают в роли системообразующих факторов становления функциональных систем разного уровня организации как при их созревании в онтогенезе, так и в процессах индивидуального обучения.
Наиболее наглядно изложенную мысль подтверждают наблюдения Е.М.Богомоловой и Ю.А.Курочкина за становлением вертикальной позы у новорожденного лосенка [7]. Первые попытки лосенка принять вертикальную позу - неуверенные и требуют значительного времени. Однако после того, как лосенок достигает потребного результата и занимает вертикальную позу, повторные его вставания при сформировавшейся функциональной системе осуществляются уверенно и быстро, в короткие отрезки времени. Под влиянием полезных приспособительных результатов (подкрепления) в соответствующих структурах центральной нервной системы функциональных систем формируется аппарат предвидения свойств потребного результата и способов его достижения - акцептор результата действия. Поэтому при возникновении той или иной потребности субъекты начинают предвидеть свойства потребного результата. В соответствующих функциональных системах формируются специальные программы поведения. Вследствие этого поведение приобретает опережающий действительные события характер и становится направленным на будущие потребные результаты.
В последние годы мы расширили понятие LсистемогенезҰ, распространив его на процессы становления, усовершенствования и угасания отдельных LсистемоквантовҰ поведения в процессе взаимодействия организма с внешней средой. Этот раздел системогенеза обозначен нами как Lсистемо-генез поведенческих актовҰ [14].
Другая кибернетическая закономерность, воплощенная в деятельность функциональных систем, - принцип саморегуляции. При этом отклонение результата деятельности любой функциональной системы от уровня, обеспечивающего нормальную жизнедеятельность, само является причиной мобилизации компонентов функциональной системы на возвращение этого результата к исходному, оптимальному для организма уровню.
Функциональные системы благодаря саморегуляции являются LквазизакрытымиҰ системами [9].
В соответствии с квантовой механикой, частицы и волны представляют собой вещество и протяженность процесса во времени. Функциональные системы и организуемые ими LсистемоквантыҰ жизнедеятельности заключают в себе свойства частицы и волны. Скорости процессов саморегуляции, их интенсивность различны у разных функциональных систем организма, что определяет ритм волновых колебательных процессов каждой из них. Слаженное взаимодействие ритмов активности различных функциональных систем в целом организме отражает еще одно их кибернетическое свойство - их резонансные свойства. Включенные в соответствующую функциональную систему элементы, особенно работающие ритмически, в свою очередь, проявляют синхронный паттерн своей активности, взаимодействуя тем самым достижению функциональной системой полезного приспособительного результата.
Важнейшие кибернетические закономерности проявляются в информационных свойствах функциональных систем организма [15], в которых постоянно осуществляется трансформация потребности в процессы нервной и гуморальной сигнализации без потери информационного значения этой потребности; нервные и гуморальные сигнализации о потребности также без потери информационного значения вызывают специфическое возбуждение мозговых структур - мотивацию; мотивация трансформируется в поведение, которое также наполнено информационным смыслом об исходной потребности организма; наконец - процессы подкрепления несут в себе полноценную информацию об удовлетворении исходной потребности [6].
Каждая функциональная система работает в организме по торсионному информационному принципу саморегуляции. Известно, что торсионные поля, действующие в пространстве, обусловлены вращательными моментами спинов взаимодействующих частиц. Рождаясь под влиянием информации, спин направлен в одну сторону, и его крутящий момент имеет одно направление. В следующий момент спин под влиянием информации направлен в другую сторону и его крутящий момент имеет другое направление.
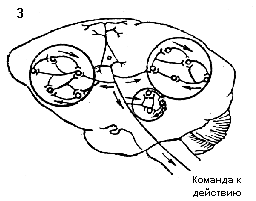
В функциональных системах организма отклонение результата деятельности функциональной системы от уровня, определяющего нормальную жизнедеятельность, заставляет все элементы функциональной системы работать в сторону его возвращения к оптимальному уровню. При этом формируется субъективный сигнал - отрицательная эмоция, позволяющая живым организмам надежно оценивать возникшую потребность. Для возвращения результата к оптимальному для жизнедеятельности уровню исполнительные элементы функциональных систем работают в противоположном направлении. Достижение оптимального уровня результата в норме сопровождается положительной эмоцией (рис. 3).
В зависимости от степени достижения требуемого результата функциональные системы усиливают или, наоборот, снижают интенсивность своей саморегуляторной деятельности. Все это определяет временные рамки основных биоритмов организма. Таким образом, внутри каждой функциональной системы постоянно действуют две противоположно направленных тенденции. Одна из них проявляется при возрастании значения результата; другая - при его снижении. Первая определяет снижение значения результата до нормального уровня, другая - его возрастание. При этом одни и те же исполнительные механизмы функциональных систем могут действовать в противоположных направлениях.Отрицательная эмоция Положительная эмоция
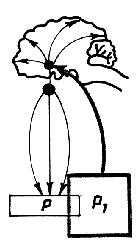
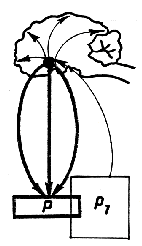
Рис.3. Торсионный механизм
деятельности функциональной системы. Слева - отклонение результата
деятельности функциональной системы от уровня, определяющего нормальный
метаболизм, активирует процессы, направленные на возвращение этого
показателя к оптимальному для жизнедеятельности уровню. Справа
- возвращение результата к исходному уровню.
Аналогичные свойства проявляются в LсистемоквантахҰ поведения. При возникновении соответствующей потребности все компоненты организма, объединенные доминирующей мотивацией, начинают мобилизовывать его на поиск веществ, удовлетворяющих исходную потребность. При достижении потребного результата мотивация исчезает и деятельность снижается. При этом субъекты нередко испытывают успокоение и даже погружаются в состояние сна. Исходная потребность и мотивация сопровождаются отрицательной эмоцией. Достижение результата (подкрепления) сопровождается положительной эмоцией, которая выступает в роли своеобразной LнаградыҰ для субъекта за усилия по достижению потребного результата.
Информационные процессы в функциональных системах организма строятся на голографическом принципе. По аналогии с физической голограммой потребность выступает в качестве опорной волны, формируя на основе мотивации аппарат предвидения потребного результата, - акцептор результата действия. С другой стороны, многоканальная обратная афферентация, поступающая к акцептору результата действия, выступает в качестве предметной волны (рис. 4).
Информационные волны о потребности и ее удовлетворении распространяются к структурам мозга в определенной временной последовательности. Все это создает условия для их интерференции и построения голограмм.
Отражение информации о метаболических потребностях и их удовлетворении в функциональных системах может осуществляться на нескольких уровнях, каждый из которых представлен своим информационным экраном.
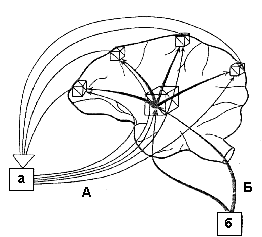
Рис.4. Схема демонстрирует
голографический механизм взаимодействия мотивации и подкрепления
на структурах мозга: а - результат поведения, б
- метаболическая потребность. А - Lпредметная сигнализация о параметрах
результатаҰ, Б - LопорнаяҰ нервная и гуморальная сигнализация
о потребности.
На клеточном уровне каждая клетка, получившая молекулярный информационный сигнал, через систему вторичных посредников и соответствующих ферментов, в частности, протеинкиназ, доводит его до генетического аппарата ядра клетки. Молекулы ДНК и РНК, будучи жидкими кристаллами, выступают в качестве Lпервого эшелонаҰ голографического информационного экрана. Под влиянием информационных сигналов о потребности они экспрессируют специальные эффекторные белковые молекулы, определяющие процессы роста, дифференцировки клеток и их метаболические реакции, опережающе направленные на устранение возникшей потребности.
Второй уровень - межклеточный информационный экран соединительной ткани, возбуждаемой многочисленными эффекторными информационными молекулами клеток тканей.
Роль голографического экрана соединительной ткани могут играть структурно организованные коллоиды межклеточного вещества - мицеллы.
Коллоиды межклеточного вещества соединительной ткани: многочисленные протеингликаны, особенно гиалуроновая кислота, а также белковые молекулы, гормоны, олигопептиды, простагландины, гликопротеины, витамины, иммунные комплексы и другие биологически активные информационные молекулы - выступают в качестве соединительнотканного LэшелонаҰ информационного экрана организма.
Поскольку протеингликаны соединительной ткани тесно взаимодействуют с гликокаликсом мембран клеток, устанавливаются тесные связи голографического экрана соединительной ткани и взаимодействующими с ней клетками. В результате формируется единый информационный экран внутренней среды организма, отражающий разнообразные стороны деятельности различных составляющих его функциональных систем.
Третий LэшелонҰ голографического экрана - структуры мозга. На этом уровне информационный голографический экран представлен коллоидами глии и молекулами ДНК и РНК, отдельных нейронов. Мозг в своей деятельности, благодаря информационным сигналам о потребностях и их удовлетворении с помощью специальных молекул, постоянно строит информационные модели действительности. Примером таких информационных моделей являются осязательная, температурная схемы, схема состояния мышечного аппарата и внутренних органов, а также - динамические стереотипы окружающей субъекта среды, которые на основе первичных процессов афферентного синтеза организуются на уровне лимбических структур и коры мозга.
Процессы афферентного синтеза, в свою очередь, активируют в голографических экранах мозга опережающие действительные события энграммы акцептора результатов действия, которые постоянно контролируют как разнообразные потребности организма, так и их удовлетворение.
Нетрудно заметить, что в информационных экранах всех уровней системной организации при наличии в организме той или иной потребности наблюдается опережающее отражение свойств потребных результатов.
Опережающее отражение действительности, открытое П.К.Анохиным [4], является универсальным свойством живого. На клеточном уровне эти процессы опережающего отражения определяются быстрыми ферментативными реакциями. Соединительная ткань за счет быстрых ферментативных реакций тоже функционирует опережающе в плане устранения различных метаболических потребностей организма, особенно при изменении рН, осмотического давления и других жизненно важных показателей внутренней среды организма. В мозговых структурах опережающие реакции определяются деятельностью акцептора результата действия. Опережающее программирование по отношению к метаболическим и гомеостатическим результатам осуществляется на основе врожденных механизмов, а по отношению к результатам поведения - на основе механизмов обучения. Наиболее выражено свойство опережающего отражения действительности у нервных клеток. На уровне периферической и особенно центральной нервной системы сложился организованный, опережающий действительные события аппарат - акцептор результата действия.
Аппарат акцептора результата действия в каждой, аппелирующей к мозгу функциональной системе широко представлен в различных структурах мозга (рис. 5). Каждая отдельная структура мозга, включенная в архитектуру акцептора результата действия, отражает свойства того или иного параметра потребного результата - его обонятельные, слуховые, зрительные, тактильные, вкусовые и другие характеристики. Системные организации акцептора результата действия складываются под влиянием свойств подкрепления и могут динамически изменяться при изменении их параметров.
1. Распространение эффекторных возбуждений пирамидного тракта по коллатералям аксонов к вставочным нейронам различных уровней ЦНС, образующим циклические комплексы многоуровневого акцептора результата действия.
2. Обратная афферентация от различных параметров результата действия поступает к соответствующим отделам акцептора результата действия. Составляющие акцептор результата действия нейроны фиксируют свойства параметров достигнутых результатов, в результате чего формируются нейрональные энграммы подкрепления.
3. Последующее распространение эффекторных влияний пирамидных нейронов опережающе активирует нейрональные энграммы акцептора результата действия, ранее сформированные подкрепляющими воздействиями. В процессах формирования и функционирования аппарата акцептора результата действия определяющая роль принадлежит обратной афферентации, поступающей в центральную нервную систему как от воздействия на соответствующие рецепторы потребности, так и при действии на многочисленные рецепторы организма параметров результатов, удовлетворяющих исходные потребности.
Приведенный материал свидетельствует о том, что функциональные системы включают в себя кибернетические, синергетические и информационные свойства.
Эти свойства функциональных систем, отработанные
миллионами лет эволюционного развития живых существ, являются
неотъемлемым атрибутом их саморегуляторной деятельности.
1. Анохин П.К. (ред.) Проблема центра и периферии в физиологии нервной деятельности.- Горький, 1935.
2. Анохин П.К., Стреж Е. Изучение динамики высшей нервной деятельности. III. Нарушение активного выбора в результате замены безусловного стимула // Физиол.журнал СССР.- 1933.- 16,2.- 28 с.
3. Анохин П.К. Теория LрезонансаҰ в нервной деятельности и ее современная критика // Архив биол. наук.- 1937.- Т.45, 2.- 5 с.
4. Анохин П.К. Опережающее отражение действительности // Вопросы философии.- 1962.- Ұ7.- C.97.
5. Анохин П.К. Биология и нейрофизиология условного рефлекса.- М.: Медицина, 1968.
6. Анохин П.К. Психическая форма отражения действительности / В кн.: Ленинская теория отражения и современность. Наука и искусство.- София, 1969.- С.109-139.
7. Богомолова Е.М., Курочкин Ю.А. Системогенез поведенческого акта / В кн.: Функциональные системы организма.- М.: Медицина, 1987.- С.353-369.
8. Винер Н. Кибернетика или управление и связь в животном и машине.- 2-у Изд. Гос.ред. Изд. для зарубежных стран.- М.: Наука, 1983.- 340 с.
9. Гладышев Г.Н. Термодинамическая теория эволюции живых существ.- М.: Луч, 1996.- 86 с.
10. Судаков К.В. Общая теория функциональных систем.- М.: Медицина, 1984.- 224 с.
11. Судаков К.В. Квантование жизнедеятельности // Успехи соврем. биологии.- 1992.- Т.112, в.4.- С.512-527.
12. Судаков К.В. (Ред.) Кванты жизнедеятельности.- М.: Изд-во ММА им. И.М.Сеченова, 1993.- 260 с.
13. Судаков К.В. Мотивация и подкрепление в системных механизмах поведения: динамические энграммы подкрепления // Журнал Высшей нервн. деятельности им. И.П.Павлова.- 1995.- Т.45, в.4.- С. 627-637.
14. Судаков К.В. (ред.) Теория системогенеза.- М.: Горизонт, 1997.- 567 с.
15. Судаков К.В. Информационные свойства функциональных систем: теоретические аспекты // Вестник РАМН.- 1997.- Ұ12.- С.3.
16. Wiener N. Cybernetics or Control and
Communication in the Animal and Machine. The Technology Press
and John Wiley and Sons Ync. N.Y.Hermann et Cie Paris, 1948.
The article demonstrates, that organism's different functional systems representing an ofjective reality possess of cybernetic features.
The functional systems have the features of self-organization
and self-control. The main cybernetic regularities manifest in
information properties of organism's functional systems. There
is an acceptor of the action result (i.e., an instrument of an
information estimation), that provides multiple feedbacks and
control processes over set parameters. The theoreical theses found
on experimental data.
Судаков Константин Викторович,
1932 года рождения. В 1956 г. окончил 1-й Московский медицинский
институт им. Сеченова, а в 1959 г. там же аспирантуру. С 1959
г. работает в 1 ММИ им. Сеченова, заведующий кафедрой нормальной
физиологии. Директор НИИ нормальной физиологии им П. К. Анохина.
Доктор медицинских наук, профессор, академик-секретарь отделения
медико-биологических наук РАМН, член Нью-Йоркской Академии наук,
член Международной организации по исследованию мозга, член Международного
общества по электросну и электроанестезии, член-корр. Американского
психосоматического общества, президент Павловского научного общества
Северной Америки, член Международного общества по изучению высшей
нервной деятельности, почетный член Венгерского общества физиологов,
вице-президент Международного общества по профилактике эмоциональных
стрессов, заслуженный деятель науки РФ, член ряда российских и
международных академий, член и председатель ряда ученых Советов,
научных советов, проблемных комиссий, экспертных советов, президиумов,
семинаров и пр. Член редколлегии 10 отечественных и зарубежных
журналов по медицине и физиологии. Участник более 40 Международных
и 80 Всесоюзных конгрессов, конференций, симпозиумов. Опубликовал
более 15 монографий - две из них переведены и изданы в США и Италии,
24 брошюры, более 600 статей, тезисов докладов и пр. Лауреат ряда
отечественных и зарубежных научных премий, награжден многими медалями
и дипломами Академий и Университетов различных стран, орденами
и медалями. Президент регионального центра профилактики стресса.