УДК 62-598.5;57.05
Успехи нейрофизиологии и других нейронаук убеждают в том, что интегративная деятельность нервной системы, основные закономерности и принципы работы нервных центров могут быть познаны, если отдельные достижения (выяснение функционального назначения нейронов различной локализации, межнейронных связей, мембранных процессов и т. д.) анализируются разносторонне и основываются на системном подходе. Становится все более очевидным, что изучение лишь Lвглубь¦, важное и необходимое само по себе, приведет в конце концов к миру элементарных частиц. Работу нейрона, нейронной сети, как и всего живого от неживого, отличает сложность организации. Вместе с тем многочисленные новые данные подтверждают правильность положений интегративной физиологии, того взгляда, согласно которому взаимодействие процессов возбуждения и торможения в ЦНС лежит в основе контроля системных функций.
Влияние тормозящих входов на активность регулирующих центров - проблема, решение которой является одной из самых трудных задач [8]. Особое внимание этой проблеме уделял П.К.Анохин при разработке теории функциональных систем [1, 2]. Он отмечал, что именно вагус наиболее тонко подключает сердце к функциональным системам организма. Изучение феномена синусовой аритмии, описанного Людвигом в прошлом веке, показало, что ослабление эфферентной импульсации в волокнах вагуса во время вдоха (и значит, растормаживание пейсмекера - водителя ритма сердца) возникает в результате поступления тормозящих афферентных сигналов к моторным вагусным нейронам. Показано, что отдельные терминали, проявляющие иммунореактивность в отношении -аминомасляной кислоты (ГАМК), образуют прямые синаптические контакты с телами и дендритами вагусных нейронов [14]. Заключение о преобладающем значении ослабления вагусных влияний для усиления работы сердца вытекает из разностороннего фармакологического анализа влияний экстракардиальных холинергических и адренергических механизмов [19].
В настоящее время признается, что в ЦНС запрограммированные нейронные сети, в которых распространяются тормозящие и возбуждающие сигналы, обеспечивают сопряженную работу нервных центров функциональных систем в обычных условиях и повышение активности пейсмекерных нейронов под влиянием возмущающих воздействий. Активность таких ключевых нейронов находится под тоническим тормозящим контролем, главным образом, по-видимому, ГАМК-ергических локальных Lсетевых¦ нейронов [20]. Она также в значительной степени зависит от таких же по характеру влияния Lпроекционных¦ нейронов других нейронных групп, где происходят процессы обработки сенсорной информации. Важно отметить, что функции тормозных передатчиков в структурах ствола мозга могут выполнять, кроме ГАМК, также норадреналин, дофамин, глицин, ацетилхолин и другие вещества и что ГАМК-ергические синапсы обнаруживаются и на тормозящих интернейронах. Отсюда при анализе информационного процессинга в структурах, контролирующих системные функции, следует определить значение растормаживания (активного торможения процесса торможения), того феномена, который является одним из важнейших открытий в нейрофизиологии.
Обширные литературные и собственные данные, полученные в опытах с использованием чрезвычайных раздражителей и веществ нейротропного действия, позволили нам обосновать представление об особом значении механизма растормаживания пейсмекерных нейронов нервных центров функциональных систем как ключевого звена в их рефлекторной деятельности [5]. Предложенная концепция основывается на признании растормаживания общим организационным принципом деятельности нервной системы [9]. Принципиальное отличие предложенной концепции от представления о центральных звеньях рефлекса как о распространяющемся от нейрона к нейрону возбуждении (иррадиация которого в ЦНС ограничивается торможением) состоит в том, что само достижение эффекта деятельности функциональной системы может быть результатом растормаживания пейсмекера - одного из центральных звеньев системы. Функции растормаживающих выключателей выполняют потоки сенсорной и другой информации к таким нейронам. При изменении спектра информации изменяется мозаика активности пейсмекеров, происходит селективное растормаживание части пейсмекеров ствола мозга в соответствии с принципами иерархии функциональных систем. Растормаживание этих нейронов и их групп как проявление участия в рефлекторной деятельности происходит при распространении соответствующих сигналов по прямым путям и через механизмы перекрестного торможения как основные звенья интеграции.
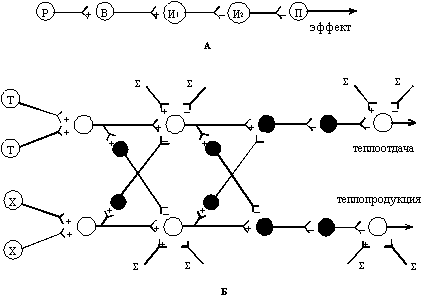
А - схематическое представление нейронного пути растормаживания пейсмекерного нейрона (П) как результата активного торможения ингибирующего нейрона И2 ингибирующим нейроном И1, который возбуждается нейроном В;
Р - рецепторный нейрон;
Б - нейронная модель центральных механизмов автономной терморегуляции, представляющая вероятные связи между путями от сенсоров (как представлено было ранее Bligh, 1984) и растормаживающими путями к пейсмекерам систем теплоотдачи и теплопродукции, обсуждаемыми в тексте: (+) - возбуждение; (-) - торможение; () - суммарные влияния.
Нетрудно представить (это следует из вышеизложенного), что особый интерес должны вызывать при использовании данного подхода центры сложных функциональных систем, в частности, системы терморегуляции. Работу таких систем позволяет понять моделирование нейронных сетей их центров. Понятно, что при применении системного подхода даже к автономной терморегуляции необходимо принимать во внимание то, что она не имеет собственных органов, а использует разные органы, ткани, их функции и разные центры. Также обращает на себя внимание то, что в технических системах нет близких аналогов из-за очевидного отсутствия потребности в них.
Попытка построения нейронной модели центрального интерфейса функциональной системы автономной терморегуляции должна быть сделана в соответствии с известными принципами ин-тегративной деятельности нервной системы [1, 2, 22]. В модели нейронных взаимодействий должны быть пред-ставлены взаимное торможение антагонистических процессов, конвергенция афферентных путей на общие эфферентные. Соответственно, главные задачи касаются связей сенсоров с эффекторами (с учетом новейших данных о тормозящем контроле тонически активных нейронов и других ключевых нейронов-осцилляторов цен-тров) и способов взаимодействия растормаживающих сигналов с возбуждающими на разных пейсмекерах ствола мозга.
Ранее было предположено, что синаптические механизмы тонического торможения вовлекаются в процессы, приводящие к повышению активности пейсмекерных нейронов, что существуют прямые связи с ними других командных центров - сенсорных миксеров как блоков аппарата сенсорной интеграции и что они могут действовать отдельно от возбуждающих путей. В соответствии с предлагаемой концепцией распространения растормаживающего возбуждения в центрах терморегуляции вероятные механизмы растормаживания пейсмекерных нейронов могут быть представлены в модели нейронных связей, как показано на рис. 1. В ней, как и в другой предложенной ранее модели [11, 12], показаны связи, выполняющие функции взаимного торможения (Lcrossing inhibition¦) разных по функциональному назначению нейронных цепей. Отличительная особенность нашей модели, как уже отмечалось [16, 17, 18], - в способе соединения сенсоров с эффекторными нейронами, которые обладают функциями пейсмекеров и посылают сигналы на периферию.
Согласно новому взгляду процессы активного торможения
процессов торможения пейсмекерных нейронов (см. рис. 1а), т.е.
процессы их растормаживания (disinhibition),
вызываются поступлением сигналов от терморецепторов. Как показано
в модели (рис. 1б), конечный эффект, т.е. изменение активности
пейсмекерного нейрона, контролирующего теплопродукцию, и пейсмекерного
нейрона, регулирующего процессы теплоотдачи, предсказуем, если
исходить из отношений путей, идущих от сенсорных нейронов (терморецепторов),
вовлекающих механизмы перекрестного торможения. Некоторые нейроны
могут интегрировать сигналы от разных терморецепторов. Нетрудно
представить, и это показано схематически (рис. 1б), что конвергенция
афферентных путей от центральных и периферических
терморецепторов к двум общим путям включает
взаимодействие уже частично интегрированных сигналов с нетемпературной
информацией. Это требует, в свою очередь, дополнительного механизма
перекрестного торможения. Последнее, таким образом, встречается
не один лишь раз на путях от сенсоров к эффекторам и оба общих
пути (Lтепловой¦ и Lхолодовой¦) участвуют в регуляции пейсмекерной
активности.
Можно представить, и это также показано в предложенной модели, что ослабление торможения некоторых пейсмекерных нейронов (например, центральных тонически активных симпатических нейронов, контролирующих теплопродукцию и сосудистый тонус), кроме растормаживания сигналами, идущими по Lхолодовым¦ путям, достигается частично в условиях действия холода и влияниями тепловых рецепторов опосредованно в результате угнетения нейронов Lтепловых¦ путей механизмами перекрестного торможения. Для таких влияний наиболее подходящим будет их определение как Lослабленное торможение¦ (Lreduced inhibition¦). Другие по функции пейсмекерные нейроны, а именно - центральные нейроны-осцилляторы, регулирующие интенсивность теплоотдачи (например, инспираторные нейроны) могут быть расторможены поступлением информационных потоков по Lтепловым¦ путям. Эти нейроны также дополнительно растормаживаются механизмом Lreduced inhibition¦, т. е. опосредованно через пути перекрестного торможения. С представлением о такой системе внутрицентральных отношений вполне согласуются известные сведения о том, что эффективные тормозные системы имеются в центрах регуляции сосудистого тонуса и перераспределения кровотока [23], ритма дыхания [15] и дрожи [13].
Таким образом, отличительная особенность модели связей между терморецепторами и центральными эффекторными нейронами (пейсмекерами и клетками с пейсмекероподобной активностью) состоит в том, что растормаживание может действовать не конвергентно, а может быть включено в качестве важнейшего регулирующего звена в эти пути (действие Lon-line¦). Иными словами, тормозящий синаптический вход - как один из путей, которые влияют на активность пейсмекера, - может быть использован для повышения его активности и это, по-видимому, так и происходит при изменении температурных условий, вовлекающих нейрофизиологические процессы, приводящие к активному торможению этого пути. Сигналы из нейронных групп-регуляторов других функциональных систем могут действовать латерально (Loff-line¦) на процессы Lафферентного синтеза¦ (по П.К.Анохину [1, 2) и непосредственно на пейсмекеры (как на рис. 1б) как Lобщие конечные пути¦ (по Ч.Шеррингтону [22).
Логика рассуждений о значении растормаживания в рефлекторной
деятельности закономерно приводит к рассмотрению вопроса о взаимодействии
между растормаживающими и возбуждающими процессами в центрах в
разных условиях. Для центров анализируемой функциональной системы
это изменения внешней температуры и температуры тела. Последние
включают сдвиги, вызываемые нейротропным действием различных по
природе химических соединений. Понятно также, что изменения в
процессах теплообмена и, следовательно, топики температур должны
вызывать в условиях действия этих факторов существенные сдвиги
в сенсорных потоках от Lядра¦ и Lоболочки¦ тела к центрам.
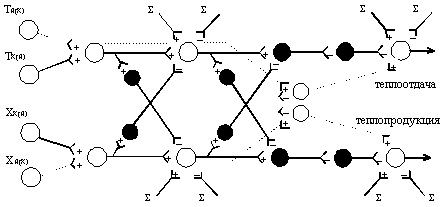
Рис. 2. Нейронная модель центральных механизмов терморегуляции у млекопитающих, включающая пути от тепловых (Т) и холодовых (Х) рецепторов кожи (к) и Lядра¦ (я) к эффекторам систем теплоотдачи и теплопродукции и растормаживающие (-) и возбуждающие (. . . .) пути к пейсмекерам, обсуждаемые в тексте:
я(к) - преобладающие влияния с терморецепторов ядра тела
к(я) - преобладающие влияния с кожных терморецепторов
(+) - возбуждение; (-) - торможение; () - суммарные влияния.
В соответствующей этим представлениям нейронной модели, очевидно, следует отразить эти отношения и, что важно, связи между терморецепторами различной локализации и растормаживающими и возбуждающими путями. Если допустить, что сигналы путей, идущих от Lядра¦ и Lоболочки¦, могут действовать на исполнительные пейсмекерные нейроны независимо друг от друга и что для некоторых центров растормаживающие пути начинаются, по-видимому, главным образом в поверхностных тканях, то модель может быть такой, как на рис. 2.
Как показано на схеме, растормаживающие и возбуждающие пути, начинаясь из разных поверхностных и глубоких тканей, могут действовать на пейсмекеры отдельно друг от друга. В настоящее время не представляется возможным представить в схеме реализацию в работе системы принципа Lопережения¦, указать, как именно в отдельных звеньях сети действуют известные тормозные и возбуждающие передатчики. Их участие в деятельности центров установлено [3, 4]. В схеме не отражено, как разные центры, содержащие пейсмекеры, могут работать иерархически и последовательно в соответствии с интенсивностью температурного и других стимулов. Вместе с тем даже такая модель позволяет понять сложность принятия решения в случаях конфликтных ситуаций в деятельности всей функциональной системы. Это может быть тогда, например, когда одна часть (половина) тела нагревается, а другая охлаждается и это приводит к поступлению в центры двух потоков информации, требующих мобилизации на периферии противоположных друг другу механизмов и процессов теплообмена. Далее можно представить, что в некоторых случаях возбуждение должно быть достаточно сильным, чтобы преодолеть тормозящие влияния на интернейроны (оказываемые из других систем) вдоль путей от сенсоров к эффекторам.
Если представить, что растормаживающие и возбуждающие пути связаны с различными рецепторами или (что легче допустить) в этих путях в разных соотношениях представлены афферентные сигналы от внешних и внутренних терморецепторов, то при этом необходимо решить вопрос о взаимодействии этих сигналов после стадий сенсорной интеграции на пейсмекерах (как общих конечных путях).
Уже в те годы, когда интенсивно изучались афферентные пути температурной чувствительности, накопились экспериментальные данные, свидетельствующие о том, что от однотипных рецепторов (например, холодовых), но различной локализации, сигналы поступают к различным нейронам гипоталамуса [21]. Установлено, что информация от поверхностных и глубоких кожных терморецепторов поступает к различным нейронам этой области [7]. В настоящее время процессы интеграции сигналов, включающие обработку информации от центральных термочувствительных нейронов, оценку средней температуры тела и ее сравнение с Lустановочной точкой¦, невозможно представить в какой-либо нейронной модели. Кроме разнообразия интегрирующих элементов следует учитывать, что соответствующие процессы включают в работу сети клетки с длительными латентными периодами (30 с и более), что может быть проявлением процессов, относящихся к накоплению сенсорной информации и растормаживанию части таких нейронов. Как показали исследования, проведенные в нашей лаборатории на срезах преоптической области переднего гипоталамуса (В.Василенко и Т.Петручук) некоторые термочувствительные ней-роны тормозятся ГАМК и ГАМКВ агонистом баклофеном. Однако поскольку ГАМКА и ГАМКВ антагонисты не оказывали заметного влияния на такие нейроны, то эти данные не позволяют считать, что они находятся под тормозящим тоническим ГАМК-ергическим контролем.
Безусловно, для разных нейронов, представляющих Lобщие конечные пути¦ и контролирующих периферические процессы термогенеза и теплоотдачи, соотношение растормаживающих и возбуждающих входов не может быть одинаковым, и при изменении температурных ус-ловий оно также изменяет-ся. Очевидно, при этом происходят закономерно сдвиги в системе взаимодействующих запрограммированных центров (и их пейсмекеров). Можно предположить, что стереотипный сдвиг возникает в условиях действия факторов, вызывающих развитие лихорадочной и подобных ей реакций. Действие холода или тепла при состояниях, развивающихся под влиянием пирогенов (включая толерантность к ним) и огромного числа фармакологических веществ, раскрывает новую сложившуюся интеграцию сигналов на нейронах. Система взаимодействующих (или точнее сказать взаимосодействующих) в поддержании температурного гомеостаза центров уже Lнастроена¦ по-другому, и при лихорадочных состояниях это может быть результатом преимущественно действия пирогенов.
Если рассматривать проблему так, что растормаживание проявляется повышением активности, т. е. пусковым действием (другое проявление - повышение возбудимости), то можно далее предположить, что функция растормаживания пейсмекеров, как эффективный пермиссивный механизм, может срабатывать и при отсутствии влияний возбуждающих путей. По-видимому, это может происходить при действии фактора, способного вызвать растормаживание пейсмекера (пусковая сигнализация) в тот момент, когда возбуждающие входы заблокированы (результат действия определенных химических соединений, например, антагонистов возбуждающих аминокислот).
Развиваемое представление о месте и роли растормаживающих сигналов в нейронных системах высших автономных центров позволяет объяснить разные проявления центрального действия синаптически активных веществ на процессы теплообмена и температуру тела. Эти эффекты известны из весьма обширной литературы и рассматривались ранее в обзорных работах [3, 4]. Их углубленный анализ с учетом нейрохимических и фармакологических аспектов проблемы приводит к необходимости внесения существенных коррективов в трактовку некоторых данных. Так, отмечалось, что гипертермический эффект вводимых в желудочки мозга простагландинов группы Е может быть Lискусственной лихорадкой¦ - результатом растормаживания ими некоторых центральных тонически активных симпатических нейронов [4, 6]. Эти нейроны, как предполагается, находятся под тормозным контролем норадренергических нейронов, а функции отрицательной обратной связи выполняют простагландины группы Е. Если это так, то нетрудно представить растормаживание этих центральных исполнительных нейронов как эффект угнетения процесса высвобождения норадреналина простагландинами, которые вводятся в Lнефизиологических¦ дозах.
Согласно развиваемому представлению, процессы интеграции сигналов приводят в зависимости от особенностей температурной и другой информации к селективному растормаживанию пейсмекеров ствола мозга в различных комбинациях и временной последовательности. Принципиально важным является то положение, что в термонейтральных условиях при отсутствии также других возбуждающих воздействий (стрессоров) растормаживающие (как и возбуждающие) влияния на пейсмекеры минимальны. Есть основания полагать, что тонические тормозящие влияния могут оказывать на пейсмекеры другие нейроны-осцилляторы.
Как отмечалось нами ранее [5, 18], контролируемые этими влияниями процессы являются по функциональному назначению трофическими и они служат цели подготовки пейсмекеров к повышению активности. В соответствии с заложенной природой программой, требующей при совершении работы соблюдение принципа экономии, такое изменение активности пейсмекеров должно развиваться вначале преимущественно под влиянием сигналов, идущих по растормаживающим путям. Применительно к рассматриваемой в данной работе более узкой проблеме центров системной регуляции процессов теплообмена можно предположить, что при последовательном изменении кожной, а затем внутренней температуры, возрастает влияние возбуждающих входов. Такая последовательность включения входов соответствует принципам работы всей системы Lпо возмущению¦ и Lпо отклонению¦.
Применение современного концептуального подхода (теории функциональных систем), конкретно - принципа последовательного динамического вовлечения элементов из конструкции связей приводит также к признанию существования достаточно определенных различий между пейсмекерами в использовании общих входов. Это очевидно так, если принять во внимание, что возбуждение таких центров системы терморегуляции, какими являются центры дрожи и потовыделения, возникает при температурных (в комбинации с другими) изменениях, характерных для развития стресс-реакции.
Таким образом, тормозящие пути, выполняющие трофические
функции, при поступлении по сенсорным каналам температурной информации,
требующей повышения активности пейсмекера, могут быть использованы
как растормаживающие выключатели. Влияния из регулирующих центров
других функциональных систем, осуществляемые конвергентно, обеспечивают
сопряжение функциональных систем и использование ими одних и тех
же элементов из конструкции связей. Дальнейшие исследования, вытекающие
из признания целесообразности развития этого представления, позволят
получить данные, важные для понимания клеточных и синаптических
центральных механизмов, относящихся к терморегуляции и другим
системным функциям организма.
1. Анохин П.К. Системные механизмы высшей нервной деятельности.- М.: Наука, 1979.
2. Анохин П.К. Биология и нейрофизиология условного рефлекса.- М.: Медицина, 1968.
3. Гурин В.Н. Центральные механизмы терморегуляции.- Минск: Беларусь, 1980.
4. Гурин В.Н. Механизмы лихорадки.- Минск: Наука и техника, 1993.
5. Гурин В.Н., Гурин А.В. Роль процессов растормаживания пейсмекеров в рефлекторной деятельности нервных центров // Физиология человека.- 1997.- Т. 23.- С. 147-149.
6. Гурин В.Н., Висмонт Ф.И., Царюк В.В. Роль центральных альфа-адренорецепторов в механизме гипертермического действия ПГЕ2 // Фармакология и токсикология.- 1984, ¦ 6.- С. 29-32.
7. Иванов К.П. Принцип энергетики организма: теоретические и практические аспекты.- Л.: Наука, 1990, Т. 1.
8. Павлов И.П. Лекции о работе больших полушарий. Полное собр. соч. в 6 т.- М.: Изд-во АН СССР, 1951, Т. 4.
9. Серков Ф.Н. Растормаживание и его роль в деятельности центральной нервной системы // Физиол. журнал СССР.- 1991.- Т. 37.- С. 107-115.
10. Судаков К.В. Основные принципы общей теории функциональных систем / В кн.: Функциональные системы организма (под ред. К.В.Судакова).- М.: Медицина, 1987.- С. 26-48.
11. Bligh J. Neuronal models of mammalian temperature regulation. In: Essays on temperature regulation. Eds. J.Blign, R.J.Moore. North-Holland, Amsterdam, 1972. P. 105-120.
12. Bligh J. Temperature regulation: a theoretical consideration incorporating Sherringtonian principles of central neurology // J. Therm. Biol. 1984. ¦ 9. P. 3-6.
13. Brck K., Hinckel P. Thermal afferents to the hypothalamus and thermal adaptation // J. Therm. Biol. 1984. ¦ 9. P. 7-10.
Simon E., Pierau F.-K., Taylor D.C. Central and peripheral thermal control of effectors in homeothermic temperature regulation. Physiol. Rev., 1986, vol.66.- P.235-300.
Sherrington C.S. The integrative action of the nervous system. Yale, New Haven, Conn., 1906.
Spyer K.M. Central nervous
mechanisms contributing to cardiovascular control. J. Physiol.,
1994, vol.474.- P.1-19.
In accordance with the principles of the theory
of functional systems and the integrative action of the nervous
system, a concept of speading of disinhibitory excitation in
the thermoregulatory centres has been developed. According to
the new idea disinhibition of pacemakers is induced by incoming
signals from thermoreceptors. A corresponding neuronal model includes
neurons capable of integrating signals coming from neuronal groups
- regulators of other functional systems, and which are able to
act convergently (Loff-line¦) on the processes of afferent synthesis
(as by P.K.Anokhin) and directly on pacemakers as Lcommon final
pathways¦ (C.Sherrington). The model also shows putative relations
between thermoreceptors of different localisation and disinhibitory
and excitatory pathways. The model also shows putative relations
between thermoreceptors of different localisation and disinhibitory
and excitatory pathways. The processes of signal integration,
depending on features of temperature and other information, lead
to selective disinhibition of brainstem pacemakers in different
combinations and temporal sequence. Under thermoneutral conditions,
in the absence of other atrressor stimuli, disinhibitory and excitatory
influences are minimal, the processes controlled by inhibitory
influences being trophic by their functional purpose and aimed
at preparing pacemakers for increased activity.