УДК 612.112+535.379:538.569
Из литературных данных известно, что действие электромагнитного излучения крайневысоких частот (ЭМИ КВЧ) приводит к модуляции иммунных реакций у человека и животных [1-3]. В медицинских исследованиях показана высокая эффективность действия ЭМИ КВЧ на организм с нарушенным функциональным состоянием [4]. В работе [5] отмечено, что биологические системы, подвергшиеся стрессу, более чувствительны к действию ЭМИ КВЧ. С позиции информационной теории взаимодействия ЭМИ КВЧ с биологическими системами [6] влияние ЭМИ КВЧ низкой интенсивности на нормально работающую клетку не существенно; в том случае, если функционирование клетки нарушено, даже слабые внешние воздействия могут менять ее метаболический профиль. В наших работах обсуждалось, что для эффекта ЭМИ КВЧ может быть существенен физиологический статус клетки [7, 8]. Возможно, что иммуномодулирующее действие ЭМИ КВЧ при патологиях направлено именно на клетки с измененным функциональным статусом. Однако специальных исследований о связи эффекта ЭМИ КВЧ и функционального состояния иммунных клеток не проводилось.
Нейтрофилы обеспечивают наиболее быстрые реакции
иммунной системы на повреждающие воздействия [9]. В организме
нейтрофилы могут находиться в покоящемся, праймированном или
активированном состояниях [10]. Нейтрофил в активированном состоянии
выполняет свою физиологическую функцию, направленную на защиту
организма от поверждающих факторов. Функциональный статус праймированного
нейтрофила характеризуется тем, что в клетке происходят метаболические
перестройки, но при этом функциональная активность не меняется,
однако при действии последующего активирующего стимула ответ клеток
многократно усиливается [11]. Агенты различной природы, в том
числе и продукты метаболизма самих нейтрофилов, могут оказывать
как праймирующее, так и активирующее действие. Изменение метаболизма
клетки при совместном действии определенных активирующих агентов
также приводит к многократному усилению ответа по сравнению с
действием отдельного агента (синергия) [12]. Мы предположили,
что реакция нейтрофилов на действие ЭМИ КВЧ с фиксированными параметрами
(частота, мощность) будет зависеть от их функционального статуса.
В работе исследовано действие ЭМИ КВЧ на нейтрофилы в различных
функциональных состояниях (интактные, праймированные и активированные).
2.1. Материалы. Использованы реактивы: кальциевый ионофор А23187, форбол 12-миристат 13-ацетат (ФМА), флуоресцентный краситель Fura-2AM, дигитонин, хемотаксический пептид N-формил-метионил-лейцил-фенилаланин (ФМЛФ), HEPES, пероксидаза хрена (тип VI), 4-бромофенацил бромид, среда 199, ЭГТА (все производства "Sigma", США), зимозан ("Биолар", СССР).
2.2. Биологический объект. Перитонеальные вызванные нейтрофилы мышей линии NMRI изолировали через 5 часов после внутрибрюшинной инъекции 150 мкл суспензии зимозана (5 мг/мл) по стандартной методике [13]. В суспензии содержалось более 85% полиморфноядерных лейкоцитов (нейтрофилов), жизнеспособность клеток составляла более 95%. Выделенные клетки хранили при +4оС в среде 199 с постоянным перемешиванием или при температуре тающего льда в солевой среде без добавления кальция. Клетки использовали через 1 час после выделения. Перед началом опыта суспензию клеток распределяли по экспериментальным ячейкам с рабочим объемом 200 мкл, плотность составляла 106 клеток/мл.
2.3. Измерение хемилюминесценции. Функциональную активность нейтрофилов оценивали по продукции активных форм кислорода (АФК) методом хемилюминесцентного анализа [14]. Хемилюминесценцию (ХЛ) измеряли, как описано в [15]. Инкубационная среда в экспериментах с ФМЛФ содержала NaN3 0,1 мМ и пероксидазу 300 ед/мл. В бескальциевый раствор для регистрации добавляли 2 мМ этиленгликольтетраацетат (ЭГТА). ХЛ регистрировали при 37оС последовательно от 12 ячеек, время опроса которых составляло 10 с в экспериментах с А23187 и ФМА или 5 с при работе с ФМЛФ.
2.4. Определение внутриклеточной концентрации свободного кальция ([Ca2+]i). Уровень [Ca2+]i измеряли с помощью флуоресцентного зонда Fura-2AM [16]. Нейтрофилы инкубировали с Fura-2AM 4 мкМ в течение 40 минут при 37оС. Затем отмывали от красителя и измеряли флуоресценцию при плотности 106 клеток/мл ( = 337 нм для возбуждения флуоресценции и = 510 нм для регистрации). [Ca2+]i рассчитывали по формуле: [Ca2+]i = Kd(F - Fmin)/(Fmax - F), где F - наблюдаемая интенсивность флуоресценции, Fmax - флуоресценция красителя, насыщенного Са2+, Fmin - флуоресценция красителя, свободного от Са2+, Kd = 224 нМ при 37оС. Fmax измеряли, добавляя 50 мкМ дигитонина к клеткам, нагруженным Fura-2AM. Собственное свечение ионофора было оценено в среде без клеток и учитывалось при расчетах [Ca2+]i.
2.5. Облучение нейтрофилов. Для облучения нейтрофилов широкополосный желобковый излучатель с апертурой 17,512,5 мм соединяли с высокочастотным генератором сигналов Г4-141 (СССР). Расстояние от излучателя до объекта устанавливали равным 400 мм, что соответствовало дальней зоне используемой антенны [15]. Клетки облучали в режиме непрерывной генерации с частотой 41,95 ГГц, плотность потока поглощенной энергии в дальней зоне антенны составляла 150 мкВт/см2. Облучение проводили при комнатной температуре 19-22оС в течение 20 мин. Одновременно облучали две пробы: (1) интактные и праймированные, или активированные клетки; (2) клетки в присутствии праймирующего, или активирующего агента, и то же с добавлением блокатора. Для каждой пробы, состоящей из 3-4 ячеек, имели соответствующий контроль необлученных клеток. После прекращения облучения регистрировали начальный уровень ХЛ, затем во все ячейки добавляли активирующий агент и продолжали регистрацию ХЛ в течение необходимого времени.
2.6. Обработка результатов.
В каждой пробе суммарную продукцию АФК оценивали по площади под
кривой зависимости ХЛ от времени (рис. 1А, 2А). Коэффициент
синергизма Кs рассчитывали как отношение усредненной
продукции АФК клеток, обработанных кальциевым ионофором А23187,
и без ионофора. Статистический анализ данных проводили с использованием
t-критерия Стьюдента. Достоверность результатов оценивали
по границам доверительного интервала с вероятностью p > 0,95.
3.1. Интактные клетки. Известные методы изоляции клеток, как и примененный нами, позволяют получать клетки в праймированном (в большей или меньшей степени) функциональном состоянии. Сразу после выделения мы наблюдали повышенные уровень спонтанной ХЛ и ХЛ в ответ на ФМЛФ (данные не приводятся). При хранении клеток в условиях низкой температуры или в бескальциевой среде функциональные показатели понижаются, что может быть связано с процессом "депраймирования" [17], и сохраняются на одном уровне в течение эксперимента. Такие клетки мы считали интактными, их физиологическое состояние принимали за исходное. После облучения ЭМИ КВЧ интактных клеток их реакция на стимулирующие агенты разной природы: кальциевый ионофор А23187, повышающий уровень [Ca2+]i, форболовый эфир, активирующий протеинкиназу С (данные не приводятся) и рецепторный агонист ФМЛФ - не изменялась. Как видно из рис.1, при облучении интактных нейтрофилов продукция АФК в ответ на ФМЛФ 1 мкМ достоверно не отличалась от контроля (611%, n = 22). Многие авторы [18] отмечали, что не подвергшиеся предварительному воздействию иммунные клетки нечувствительны к действию низкочастотных ЭМП. Авторы работы [3] обращают внимание на то, что определенные функциональные показатели иммунных клеток, выделенных из облученных ЭМИ КВЧ мышей с нормальным иммунитетом, не отличались от контроля. Возможно, это является общим свойством биологических систем во взаимодействии с низкоинтенсивным излучением.
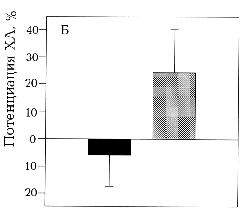
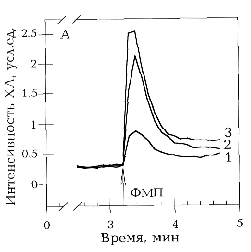
А - оригинальные записи зависимости интенсивности
ХЛ от времени при активации хемотаксическим пептидом ФМЛФ 1 мкМ
(1) интактных нейтрофилов, (2) праймированных и (3) облученных
ЭМИ КВЧ праймированных нейтрофилов. Время подачи активирующего
стимула указано стрелкой. Б - эффект ЭМИ КВЧ на интактные () и
праймированные () нейтрофилы, рассчитанный по отношению продукции
АФК за определенный период облученных и необлученных нейтрофилов.
Для праймирования использован ФМЛФ 10 нМ за 20 минут до активации.
Клетки облучали ЭМИ КВЧ в режиме непрерывной генерации с частотой
41.95 ГГц в дальней зоне антенны при плотности потока поглощенной
энергии 150 мкВт/см2 в течение 20 минут при комнатной
температуре (то же для рис. 2 и 3). Указан доверительный
интервал р>0,95, рассчитанный по критерию Стьюдента.
3.2. Праймированные клетки.
Праймирование клеток ФМЛФ в низкой концентрации 1 нМ подготавливало
их к более сильной реакции на последующую активацию ФМЛФ в высокой
концентрации 1 мкМ (рис.1А). Мы проверили, будет ли эффективным
действие ЭМИ КВЧ на праймированные нейтрофилы. Вследствие облучения
праймированных нейтрофилов продукция АФК достоверно возрастала
на 24,315,9% (n = 22) по сравнению с необлученными
клетками. Механизм праймирования в настоящее время остается во
многом неопределенным. Считается, что праймирование может быть
результатом взаимодействия Са2+-зависимых сигнальных
систем, действующих через серин/треониновые киназы, и сигнальных
систем тирозиновых киназ [11]. Вероятно, ЭМИ КВЧ может влиять
на клетки на уровне взаимодействия киназ, что приводит к значительному
потенцированию их ответа на активирующий агент. Возможно, что
вызванное праймированием изменение активности определенных киназ
повышает чувствительность клеток к ЭМИ КВЧ.
3.3. Активированные клетки. Добавление кальциевого ионофора А23187 в среду инкубации приводило к повышению уровней ХЛ и [Ca2+]i, что определяло исходный уровень активации клеток (данные не приводятся). Мы проводили облучение нейтрофилов, предварительно обработанных А23187 в концентрациях от 0-,120 мкМ, по окончании облучения клетки активировали ФМА 1 мкМ. На рис. 2А показан типичный пример синергического ответа нейтрофилов на совместное действие А23187 и ФМА и изменение ответа вследствие действия ЭМИ КВЧ. Эффект облучения зависел от наличия Са2+ в среде и концентрации ионофора: в бескальциевой среде во всем диапазоне концентраций А23187 ответ на ФМА облученных и необлученных клеток не различался; в среде, содержащей Са2+, при возрастании концентрации ионофора наблюдалось увеличение эффекта ЭМИ КВЧ (рис. 2Б). Возможно, такая зависимость определяется уровнем [Ca2+]i. Известно, что кальциевые ионофоры вызывают повышение [Ca2+]i, высвобождая Са2+ из внутриклеточных депо в начальный момент, обеспечивая ионофорный перенос и вход Са2+ через неспецифические катионные каналы плазматической мембраны в течение всего времени действия ионофора [19, 20]. При добавлении ионофора наблюдается кратковременное значительное повышение [Ca2+]i, которое сменяется постепенным возвратом к исходному уровню или поддерживается повышенным продолжительное время при высоких концентрациях ионофора (табл.). Сопоставление результатов по измерению [Ca2+]i и эффектам ЭМИ КВЧ показывает, что достоверные различия коэффициентов синергизма облученных и необлученных клеток (рис.2Б) обнаруживаются в среде, содержащей Са2+, при высоких концентрациях ионофора, на фоне которых в клетке создается повышенный поддерживаемый уровень [Ca2+]i, что создает условия для значительной активации ферментов. Вероятно, предшествующий облучению уровень активации Ca2+-зависимых ферментов определяет метаболические перестройки нейтрофила, вызванные действием ЭМИ КВЧ.
Таблица
|
|
|
|
|
| ||
Примечание: *n - число экспериментов. Даны средние
значения n.
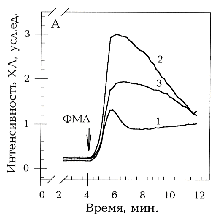
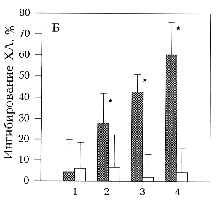
А - оригинальные записи зависимости интенсивности
ХЛ от времени при активации форболовым эфиром ФМА 1 мкМ (момент
подачи указан стрелкой) интактных нейтрофилов (1), необлученных
(2) и облученных ЭМИ КВЧ (3) нейтрофилов, предварительно обработанных
кальциевым ионофором А23187 7,5 мкМ (синергическая активация).
Б - ингибирование КВЧ-излучением синергического ответа нейтрофилов
на активацию ФМА 1 мкМ и А23187 в разных концентрациях (1
- 0,5 мкМ, 2 - 5 мкМ, 3 - 10 мкМ, 4 - 20 мкМ).
Среда инкубации содержала 1 мМ Са2+ () или 2 мМ
ЕГТА без добавления Са2+ (). Представлено процентное
отношение коэффициентов синергизма Кs облученных и
необлученных нейтрофилов. Указан доверительный интервал р>0,95,
рассчитанный по критерию Стьюдента. - Кs
облученных и необлученных нейтрофилов отличаются достоверно.
3.4. Эффект ингибирования фосфолипазы А2. Известно, что одним из наиболее активных Ca2+-зависимых ферментов в нейтрофилах, участвующих в регуляции респираторного взрыва, является фосфолипаза А2 (ФЛА2) [9], показан ее вклад в синергическую активацию нейтрофилов кальциевым ионофором и ФМА [12]. Действие А23187 в концентрациях, превышающих 0,5 мкМ, может вызывать неспецифическую активацию фосфолипаз, в том числе ФЛА2 [12]. Арахидоновая кислота, продукт ФЛА2, является агентом, потенцирующим активность протеинкиназы С (ПКС) в активации респираторного взрыва [9]. Активность ФЛА2 усиливается фосфорилированием, и ее продукт - арахидоновая кислота - стимулирует ПКС, таким образом обеспечивая положительную обратную связь, которая регулируется Са2+-зависимостью обоих ферментов. Мы исследовали роль ФЛА2 в метаболической перестройке нейтрофила при действии ЭМИ КВЧ на примере синергической реакции А23187 и ФМА. Для этого сравнивали эффекты ЭМИ КВЧ на клетки, обработанные А23187 5-8 мкМ в присутствии блокатора ФЛА2 4-бромофенацил бромида (BrPhBr) 10 мкМ и без него. По реакции на BrPhBr выделены две группы клеток (рис.3А): 1) клетки с высокой чувствительностью, у которых происходило уменьшение Ks от 2,4¦0,2 до 1,0¦0,1 при действии BrPhBr (n = 21); 2) клетки с низкой чувствительностью, Ks которых, составляющий 2,8¦0,3 на фоне ионофора, уменьшался до 2,1¦0,2 при добавлении BrPhBr (n = 15). В первой группе клеток (рис.3Б) при действии ЭМИ КВЧ наблюдалось ингибирование ХЛ на 24¦10%, добавление BrPhBr снижало эффект ЭМИ КВЧ до 7¦7%, что достоверно не отличалось от контроля. Во второй группе клеток облучение не было эффективным (рис. 3Б). Таким образом, ингибирование синергической реакции А23187 и ФМА в активации респираторного взрыва при действии ЭМИ КВЧ определялось активностью ФЛА2: эффект был значимым только в случае высокой активности фермента и отсутствовал при низкой. Полученный результат дает основание считать, что ФЛА2 является одной из мишеней действия ЭМИ КВЧ в нейтрофилах. Известно, что для полной активации ФЛА2 необходимы фосфорилирование и транслокация к мембране [21]. Транслокация зависит от Са2+-зависимого фосфолипид-связывающего домена, и наблюдается при действии агентов, увеличивающих [Са2+]i, например, кальциевого ионофора. Фосфорилирование может быть обеспечено активированной ПКС, например, при действии ФМА. Следует отметить, что транслокация и фосфорилирование независимы в регуляции активности ФЛА2 [22]. На макрофагах показано, что блокирование ФЛА2 может идти с разной эффективностью по двум указанным путям: сильно ингибируется активация через ПКС и частично - через повышение [Са2+]i [21]. Возможно, что выделенные группы клеток различаются по вкладу в регуляцию активности ФЛА2 каждого из этих путей, и действие ЭМИ КВЧ более эффективно в случае преобладания фосфорилирования, что может объясняться направленным влиянием ЭМИ КВЧ именно на процессы фосфорилирования.
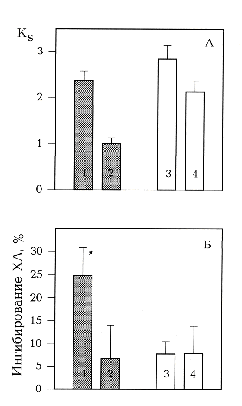
А - коэффициенты синергизма Кs в группах клеток с высокой () и низкой () чувствительностью к блокатору фосфолипазы А2 4-бромофенацил бромиду (BrPhBr) 10 мкМ. Б - действие ЭМИ КВЧ на нейтрофилы с высокой () и низкой () чувствительностью к BrPhBr 10 мкМ, определенное по отношению Кs облученных и необлученных нейтрофилов.
1, 3 - клетки в среде с А23187 5 мкМ; 2, 4 - клетки в среде с А23187 5 мкМ с добавлением BrPhBr 10 мкМ. Указан доверительный интервал р>0,95, рассчитанный по критерию Стьюдента. - Кs облученных и необлученных нейтрофилов отличаются достоверно.
Приведенные в работе экспериментальные данные показывают, что функциональный статус определяет эффект действия ЭМИ КВЧ на клетку, усиливая, ослабляя или не изменяя ее ответ на активирующий агент. В работе [18], где приведены обобщенные данные по действию низкочастотного ЭМИ на лимфоциты, отмечено, что в зависимости от степени активации клетки будет наблюдаться стимуляция, ингибирование или отсутствие эффекта. По всей вероятности, взаимодействие ЭМИ с клеткой происходит на уровне активированной ферментной системы. Процессы праймирования и активации нейтрофила имеют общие пути. Ключевыми звеньями в праймировании и активации нейтрофилов являются система Са2+-сигнализации, действующая через серин/треониновые киназы, и система сигнализации тирозиновой киназы [10, 11]. В последовательности ФЛА2 из фагоцитов представлены потенциальные сайты фосфорилирования для обоих серин/треониновых и тирозиновых киназ [9]. Возможно, что вызванные ЭМИ КВЧ метаболические перестройки в праймированных и активированных нейтрофилах могут определяться уровнем фосфорилирования определенных ферментов.
Авторы выражают глубокую признательность И.В.Байдаровой
за техническую помощь и Б.Ф.Санталову за оригинальные конструкторские
решения и их воплощение в приборном обеспечении экспериментов.
The work considers the long-standing question on
the relation between the interaction effectiveness of EHF EMF
with an immune cell and a functional status of the cell. The physiological
activity was evaluated on the respiratory burst intensity. The
cells were irradiated by continuous EHF EMF (41,95 GHz, 150 mW/cm2)
in the far field zone of the channel radiator. We have shown that:
1) intact cells did not change their response to stimuli
after EHF EMF irradiation; 2) at a change of the cell functional
status by calcium ionophore A23187 pretreatment the EHF EMF considerably
inhibited the response to subsequent activation by PMA; 3) the
EHF EMF considerably enhanced response of primed cells to subsequent
activation; 4) the sensitivity of cells to EHF EMF depended
on [Ca2+]i, 5) an inhibiting effect
of EHF EMF on neutrophil respiratory burst was significant only
if activity of phospholipase A2 was high. EHF EMF can enhance
or reduce the response to the activating agent depending on functional
status of the cell. The results obtained show that the interaction
of EHF EMF with cell occurs at the level of activated enzymes.
Аловская Алла Анатольевна окончила Нижегородский государственный университет и магистратуру Пущинского государственного университета, учится в аспирантуре того же университета. Защитила магистерскую диссертацию по действию КВЧ-излучения на дыхательный взрыв в нейтрофилах мыши. Автор 11 опубликованных научных работ. Область научных интересов: внутриклеточная сигнализация и биологические эффекты электромагнитных полей.
Габдулхакова Аида Габдрахмановна
окончила биологический факультет Казанского государственного университета
и магистратуру Пущинского государственного университета. В настоящее
время учится в аспирантуре ПущГУ. Защитила магистерскую диссертацию
по внутриклеточной сигнализации нейтрофилов и действию ЭМИ КВЧ.
Автор 10 опубликованных научных работ. Область научных интересов:
клеточная биология, внутриклеточная сигнализация, биофизика и
биохимия клетки.
Гапеев Андрей Брониславович,
1968 года рождения. Окончил в 1992 году физический факультет Казанского
государственного университета им. В.И.Ульянова-Ленина по специальности
радиофизика и электроника. С 1994 года работает младшим научным
сотрудником в Институте биофизики клетки РАН. В 1997 году защитил
диссертацию кандидата физико-математических наук. Автор 20 опубликованных
научных работ. Область научных интересов: электромагнитобиология,
внутриклеточная сигнализация.
Дедкова Елена Николаевна
окончила ветеринарно-биологический факультет Московской ветеринарной
академии им. К.И.Скрябина. В настоящее время является аспирантом
кафедры физиологии клетки Пущинского государственного университета.
Автор 4 научных статей. Область научных интересов: внутриклеточная
сигнализация, физиология и биохимия клетки.
Сафронова Валентина Григорьевна
окончила биологический факультет Томского государственного университета
и аспирантуру Института биологической физики АН СССР, кандидат
биологических наук, старший научный сотрудник лаборатории биофизики
нервной клетки Института биофизики клетки РАН. Защитила диссертацию
по регуляции электрогенеза и сократимости миокарда. Автор 45 опубликованных
научных работ. Область научных интересов: биофизика и биохимия
клетки, биологические эффекты электромагнитных полей, внутриклеточная
сигнализация.
Фесенко Евгений Евгениевич
родился в 1939 году в Ростове-на-Дону. В 1963 году окончил физико-математический
факультет Ростовского государственного университета. Дальнейшая
его трудовая и научная деятельность связана с г. Пущино. Работал
в Институте биологической физики АН, затем - директор Института
биофизики клетки РАН, доктор биологических наук, профессор, член
Европейской академии, первый заместитель председателя президиума
Пущинского научного центра, главный редактор журнала "Биофизика",
зам. председателя совета по проблеме "Биологическая физика",
председатель комиссии Международного союза чистой и прикладной
биофизики (IUPAB). Им опубликовано более 200 научных работ, из
них три работы - в журнале "Nature". Основные научные
интересы связаны с биофизикой рецепции и электромагнитобиологией.
Чемерис Николай Константинович родился в 1942 году в г. Ульяновске. Закончил кафедру биофизики Московского государственного университета, работает в Институте биофизики клетки РАН, профессор Тульского государственного педагогического университета им. Л.Н.Толстого и Пущинского государственного университета, доктор биологических наук, Лауреат государственной премии. Им опубликовано более 80 научных работ. Основные научные интересы связаны с электромагнитобиологией и внутриклеточной сигнализацией.