хДЕМРХТХЙЮЖХЪ ЪБКЕМХЪ ЯХМЕПЦХГЛЮ Б ТСМЙЖХНМЮКЭМШУ
ЯХЯРЕЛЮУ НПЦЮМХГЛЮ ВЕКНБЕЙЮ Х ФХБНРМШУ Я ОНГХЖХИ ЙНЛОЮПРЛЕМРМН-ЙКЮЯРЕПМНЦН
ОНДУНДЮ
б.л. еЯЭЙНБ*, ч.л. оНОНБ*,
м.ю. тСДХМ, ю.ю. уЮДЮПЖЕБ
бБЕДЕМХЕ. ъБКЕМХЕ ЯХМЕПЦХГЛЮ Б АХНЯХЯРЕЛЮУ
- РПЮДХЖХНММЮЪ РЕЛЮ ХЯЯКЕДНБЮМХИ Б ТХГХНКНЦХХ (Б ВЮЯРМНЯРХ, Б МЕПБМН-ЛШЬЕВМНИ
ТХГХНКНЦХХ), ПЪДЮ ЙКХМХВЕЯЙХУ ДХЯЖХОКХМ (ЯХМЕПЦХВЕЯЙХЕ
БГЮХЛННРМНЬЕМХЪ ЛЕФДС НПЦЮМЮЛХ ХКХ ЯХЯРЕЛЮЛХ НПЦЮМНБ ЯОНЯНАЯРБСЧР АНКЕЕ СЯЙНПЕММНЛС БШГДНПНБКЕМХЧ, ЯНЙПЮЫЕМХЧ ОНЯКЕНОЕПЮЖХНММНЦН
ОЕПХНДЮ Х Р.Д.).
гЮ ОНЯКЕДМХЕ ЦНДШ ХМРЕПЕЯ Й ЯХМЕПЦХВЕЯЙХЛ
ОПНЖЕЯЯЮЛ Б ОПХПНДЕ ПЕГЙН БШПНЯ ЯПЕДХ АХНКНЦНБ Х ЛЮРЕЛЮРХЙНБ. щРН НАЗЪЯМЪЕРЯЪ
ЮЙРХБМШЛ ХЯЯКЕДНБЮМХЕЛ УЮНРХВЕЯЙХУ ЯРПСЙРСП, ХЯЯКЕДНБЮМХЪЛХ Б НАКЮЯРХ ЮБРНБНКМНБШУ ОПНЖЕЯЯНБ Х ХГСВЕМХЕЛ ЖЕКНЦН ПЪДЮ АХНЯТЕПМШУ
ОПНЖЕЯЯНБ. бН БЯЕУ ЩРХУ ЯКСВЮЪУ БНГМХЙМНБЕМХЕ ОНПЪДЙЮ, ЯРПСЙРСП ХГ УЮНЯЮ ХКХ
ПНФДЕМХЕ БНКМ ЛНФЕР АШРЭ НАСЯКНБКЕМН ЯХМЕПЦХВЕЯЙХЛХ
БГЮХЛНДЕИЯРБХЪЛХ ЛЕФДС ЩКЕЛЕМРЮЛХ Б УЮНЯЕ (ХКХ Б ЮЙРХБМНИ ЯПЕДЕ).
хЯЯКЕДНБЮМХЧ
НАЫХУ БНОПНЯНБ ЯРПСЙРСПХПНБЮМХЪ Б УЮНРХВЕЯЙХУ ЯПЕДЮУ (МЮВХМЮЪ Я ЛНКЕЙСКЪПМНЦН Х
ЙНМВЮЪ ОНОСКЪЖХНММН-АХНЯТЕПМШЛ СПНБМЕЛ) ЯРЮКН БЕЯЭЛЮ ЮЙРСЮКЭМНИ ОПНАКЕЛНИ Х
КЕЦКН Б НЯМНБС ЮЙРХБМН ПЮГБХБЮЧЫЕИЯЪ ЯХМЕПЦЕРХЙХ. аЮГНБШЕ ОПНАКЕЛШ ЩРНИ МЮСЙХ ЯБЪГЮМШ Я ХДЕМРХТХЙЮЖХЕИ
БНГЛНФМНЯРЕИ ЯХМЕПЦХВЕЯЙХУ БГЮХЛННРМНЬЕМХИ ЛЕФДС
ЩКЕЛЕМРЮЛХ (ЙНЛОЮПРЛЕМРЮЛХ) ХГСВЮЕЛНИ ЯПЕДШ (НАЗЕЙРЮ)
Х, Б ВЮЯРМНЯРХ, Б АХНЯПЕДЮУ, ЙЮЙ НДМНЦН ХГ НАЗЕЙРНБ
ХГСВЕМХЪ Б ЯХМЕПЦЕРХЙЕ. хЛЕММН АХНЯПЕДШ Х АХНАЗЕЙРШ ЯНЯРЮБКЪЧР МЮХАНКЕЕ ХМРЕПЕЯМШИ НАЗЕЙР ХГСВЕМХЪ
[1, 4].
пЮЯЯЛНРПХЛ ТСМЙЖХНМЮКЭМШЕ
ЯХЯРЕЛШ НПЦЮМХГЛЮ (тян) ЛКЕЙНОХРЮЧЫХУ. яХМЕПЦХВЕЯЙХЕ
ОПНЖЕЯЯШ ОПНХЯУНДЪР Б ЙКЮЯРЕПЮУ тян (МЮОПХЛЕП, Б ЙЮПДХН-ПЕЯОХПЮРНПМНИ
ЯХЯРЕЛЕ (йпя), Б МЕИПНЯЕРЪУ ДШУЮРЕКЭМНЦН ЖЕМРПЮ (мп
дж) - ЩРН ОПНЖЕЯЯШ, Ю Б ЯЕПДЖЕ - ЮБРНБНКМНБШЕ
ЯХМУПНМХГХПНБЮММШЕ ОПНЖЕЯЯШ). яХМЕПЦХВЕЯЙХЕ ОПНЖЕЯЯШ
Б ЩРХУ ЯХЯРЕЛЮУ НОПЕДЕКЪЧР СЯРНИВХБНЯРЭ ДШУЮРЕКЭМНИ ПХРЛХЙХ Х
ОЕПХНДХВЕЯЙСЧ ПЮАНРС ЯЕПДЖЮ. щРН НАЕЯОЕВХБЮЕР ОНЯРНЪМЯРБН ЦЮГНБНЦН ЯНЯРЮБЮ
ЙПНБХ Х СЯРНИВХБСЧ ПЮАНРС НПЦЮМХГЛЮ, Р.Е. ЦНЛЕНЯРЮГ. хГСВЕМХЕ ЯХМЕПЦХГЛЮ Б йпя
- НДХМ ХГ БЮФМШУ ПЮГДЕКНБ ЯХМЕПЦЕРХЙХ, Ю ХГСВЕМХЕ ЯРЕОЕМХ ЯХМЕПЦХГЛЮ Б тян
ЛНФЕР ЯКСФХРЭ ПЮГБХРХЧ РЕНПЕРХВЕЯЙНИ ЛЕДХЖХМШ Х ТХГХНКНЦХХ РПСДЮ Х ЯОНПРЮ.
жЕКЭ ПЮАНРШ
- НЖЕМЙЮ ЯХМЕПЦХГЛЮ Б тян МЮ ОПХЛЕПЕ йпя; ЛЕРНДШ Х ЮКЦНПХРЛШ ХДЕМРХТХЙЮЖХХ
ЙНЩТТХЖХЕМРНБ ЯХМЕПЦХГЛЮ Б ЮЙРХБМШУ ЯПЕДЮУ (пмя Х ЯЕПДЕВМНИ ЛШЬЖЕ).
лЕРНДШ Х
ЮКЦНПХРЛШ ХЯЯКЕДНБЮМХЪ ЯХМЕПЦХГЛЮ Б тян. б АХНКНЦХХ ЯХМЕПЦХВЕЯЙХЕ
НРМНЬЕМХЪ НОХЯШБЮЧРЯЪ ЙЮВЕЯРБЕММН. йНКХВЕЯРБЕММШЕ УЮПЮЙРЕПХЯРХЙХ ХЛЕЧР ЯРЮРСЯ
ОПЪЛШУ ХГЛЕПЕМХИ (ЮЙРХБМНЯРЭ ЩКЕЛЕМРНБ пмя, ЯХКЮ ЛШЬЕВМНЦН ЯНЙПЮЫЕМХЪ Х Р.Д.) Х
МЕ ХЛЕЧР ЕДХМНЦН ОНДУНДЮ. мН РЮЙНИ ОНДУНД ЯСЫЕЯРБСЕР. нМ АЮГХПСЕРЯЪ МЮ НЯМНБМШУ
ОПХМЖХОЮУ ЯХМЕПЦХГЛЮ АХНКНЦХВЕЯЙХУ ДХМЮЛХВЕЯЙХУ ЯХЯРЕЛ (адя).
лШ ЛНФЕЛ БШДЕКХРЭ ЯПЮГС ДБЮ
РЮЙХУ НЯМНБМШУ ОПХМЖХОЮ. цКЮБМШИ ХГ МХУ ЯБЪГЮМ Я РПЕАНБЮМХЕЛ НРЯСРЯРБХЪ ХКХ, Б
ЙПЮИМЕЛ ЯКСВЮЕ, ЛХМХЛХГЮЖХХ РНПЛНГМН-СЦМЕРЮЧЫХУ
ЯБЪГЕИ Б адя. б ЛНДЕКЪУ адя НМХ ЛНЦСР АШРЭ ОПЕДЯРЮБКЕМШ ХКХ МЮКХВХЕЛ
НРПХЖЮРЕКЭМШУ БЕЯНБШУ ЯБЪГЕИ ЛЕФДС ЙНЛОЮПРЛЕМРЮЛХ ХКХ
ЙКЮЯРЕПЮЛХ адя, ХКХ МЮКХВХЕЛ БЕМРХКЭМШУ (САШБЮЧЫХУ) ТСМЙЖХИ, ЯРНЪЫХУ ОЕПЕД
ОЕПЕЛЕММШЛХ Б ЯХЯРЕЛЮУ ДХТТЕПЕМЖХЮКЭМШУ СПЮБМЕМХИ, ОПЕДЯРЮБКЪЧЫХУ адя Б ПЮЛЙЮУ ЙНЛОЮПРЛЕМРМН-ЙКЮЯРЕПМНЦН ОНДУНДЮ (ййо) [2-3].
б ОЕПБНЛ ЯКСВЮЕ
(НРПХЖЮРЕКЭМШЕ ЩКЕЛЕМРШ Юij ЛЮРПХЖ ЛЕФЙНЛОЮПРЛЕМРШУ
ХКХ ЛЕФЙКЮЯРЕПМШУ ЯБЪГЕИ ю) ХЛЕЧРЯЪ ЯКНФМНЯРХ Я КХМЕЮПХГЮЖХЕИ ДХМЮЛХЙХ ОНБЕДЕМХЪ адя Х
ОПХБЕДЕМХЕЛ ЛЮРПХЖШ ЯБЪГЕИ ю Й НЙНМВЮРЕКЭМН МЕНРПХЖЮРЕКЭМНЛС БХДС. бН БРНПНЛ ЯКСВЮЕ
ОПНАКЕЛЮ СЯКНФМЪЕРЯЪ ЕЫЕ АНКЭЬЕ, Р.Й. НМЮ НАСЯКНБКЕМЮ МЕНДМНГМЮВМНЯРЭЧ БХДЮ
ЛЮРПХЖШ НАПЮРМШУ ЯБЪГЕИ п(С), ОПНХГБНКЭМНЦН БШАНПЮ ЕЕ ОЮПЮЛЕРПНБ Х
БХДЮ ТСМЙЖХИ pi (С)
[2-3]. мЮИДЕМН ПЕЬЕМХЕ ДКЪ ЙНМЙПЕРМШУ ЛЕДХЙН-АХНКНЦХВЕЯЙХУ ОПХЛЕПНБ. хЛЕММН ЩРХ
ПЕГСКЭРЮРШ Х ЮКЦНПХРЛШ ХДЕМРХТХЙЮЖХХ пмя Х яяя ПЮЯЯЛНРПХЛ ОНДПНАМН.
б ПЮЛЙЮУ ОЕПБНЦН ОНДУНДЮ
МЕНАУНДХЛН СДНЯРНБЕПХРЭЯЪ Б КХМЕИМНЛ (ХКХ ЙБЮГХКХМЕИМНЛ Б АХНКНЦХВЕЯЙНЛ ЯЛШЯКЕ)
ОНБЕДЕМХХ ХЯЯКЕДСЕЛНИ адя. дКЪ ЩРНЦН БШАХПЮЧРЯЪ
ЮДЕЙБЮРМШЕ БУНДМШЕ БНГДЕИЯРБХЪ Х ХЯЯКЕДСЧРЯЪ ЯННРМНЬЕМХЪ ЛЕФДС
БУНДМШЛХ Х
БШУНДМШЛХ БЕКХВХМЮЛХ. дКЪ пмя БУНДМШЛХ БЕКХВХМЮЛХ ЪБКЪКХЯЭ ПЮГДПЮФЮЧЫХЕ
ЩКЕЙРПХВЕЯЙХЕ ХЛОСКЭЯШ, С ЙНРНПШУ ХГЛЕМЪКХЯЭ ЯХКЮ Х ДКХРЕКЭМНЯРЭ ЯРХЛСКНБ
(НАШВМН ЩРН ЯЕПХХ ЯРХЛСКНБ). дКЪ яяя ЩРН ЛНЦСР АШРЭ РНФЕ ЩКЕЙРПНЯРХЛСКШ,
МЮМНЯХЛШЕ МЮ МЕИПНЯЕРХ ЯНЯСДНДБХЦЮРЕКЭМНЦН ЖЕМРПЮ ХКХ
ДНГХПНБЮММШЕ ТХГХВЕЯЙХЕ МЮЦПСГЙХ (ОПХЯЕДЮМХЪ, ЯРЕО-РЕЯРШ),
СВЮЫЕММНЕ ДШУЮМХЕ ОПХ РЕЯРЮУ Я НЙЯХЦЕЛНЛЕРПНЛ. б ЩРНЛ
ЯКСВЮЕ МЮДН ХЛЕРЭ БНГЛНФМНЯРЭ СОПЮБКЪРЭ ОЮПЮЛЕРПЮЛХ БУНДМШУ БЕКХВХМ Х
ПЕЦХЯРПХПНБЮРЭ БЕКХВХМС БШУНДМШУ ЯХЦМЮКНБ (ЮЙРХБМНЯРЭ ЩТТЕПЕМРМШУ МЕПБНБ,
ХГЛЕМЕМХЪ ВЮЯРНРШ ЯЕПДЕВМШУ ЯНЙПЮЫЕМХИ (вяя) ХКХ ЮПРЕПХЮКЭМНЦН ДЮБКЕМХЪ (юд)).
дКЪ КХМЕИМШУ ЯХЯРЕЛ ОПХ ЩРНЛ ДНКФМШ МЮАКЧДЮРЭЯЪ НОПЕДЕКЕММШЕ ЯННРМНЬЕМХЪ ЛЕФДС
БУНДНЛ Х БШУНДНЛ Б ПЮЛЙЮУ ЛНДЕКЕИ КХМЕИМНЦН ВЕПМНЦН
ЪЫХЙЮ (квъ).
нРЛЕРХЛ ЙПЮРЙН НЯМНБМШЕ
ЯБНИЯРБЮ Х НЯНАЕММНЯРХ НПЦЮМХГЮЖХХ тян, Н ЙНРНПШУ ЛШ ЛНФЕЛ ЦНБНПХРЭ ЙЮЙ Н
ЙБЮГХКХМЕИМШУ НАЗЕЙРЮУ (квъ). еЯКХ МЮ БУНДЕ адя ГЮДЮЕРЯЪ МЕГЮБХЯХЛЮЪ ОЕПЕЛЕММЮЪ
U1(t) Х
ДКЪ МЕЕ МЮ БШУНДЕ ОНКСВЮЕРЯЪ ГЮБХЯХЛЮЪ ОЕПЕЛЕММЮЪ С1(t), Ю ГЮРЕЛ ДКЪ ДПСЦНИ
ОЕПЕЛЕММНИ U2(У) ОНКСВЮЕРЯЪ МЮ БШУНДЕ С2(t), РН
ДКЪ квъ ОПХ БУНДМНИ ОЕПЕЛЕММНИ Ю1u1(t)+Ю2U2(t) ОНКСВЮЕРЯЪ
БШУНДМЮЪ БЕКХВХМЮ
С3=Ю1С1(t)+Ю2С2(У). (1)
еЯКХ ХЛЕЕЛ Й БУНДНБ, РН
КХМЕИМЮЪ ЙНЛАХМЮЖХЪ БУНДМШУ БЕКХВХМ (ОН ЩРХЛ БУНДЮЛ) ДНКФМЮ ДЮРЭ ОПНЯРСЧ
КХМЕИМСЧ ЯСОЕПОНГХЖХЧ БШУНДМШУ БЕКХВХМ, Р.Е. С=S Ю1u1(t).
нДМНБПЕЛЕММН, ЕЯКХ С квъ ХЛЕЕРЯЪ МЕЯЙНКЭЙН БШУНДНБ, РН КХМЕИМНЕ ХГЛЕМЕМХЕ
ЯХЦМЮКЮ МЮ НДМНЛ БУНДЕ ДНКФМН КХМЕИМН (Б Ю
ПЮГ) ХГЛЕМЪРЭ БЯЕ БШУНДМШЕ ЯХЦМЮКШ С1(t). хЛЕММН РЮЙХЕ ОПНБЕПЙХ щбл Х ОПНХГБНДХР ДКЪ пмя, Б
ЙНРНПНИ ХЛЕЕРЯЪ ЛМНФЕЯРБН ЮТТЕПЕМРМШУ Х ЩТТЕПЕМРМШУ ЙЮМЮКНБ. дКЪ ЯКСВЮЪ яяя
ХЛЕЕЛ НДХМ БУНД Х НДХМ БШУНД, ОНЩРНЛС ХДЕМРХТХЙЮЖХЪ квъ МЕ БШГШБЮЕР НЯНАНЦН
РПСДЮ. оНЯКЕ ХДЕМРХТХЙЮЖХХ квъ ДКЪ йпя ЛШ ХЯОНКЭГНБЮКХ ЛЕРНД ХДЕМРХТХЙЮЖХХ
ЛЮРЕЛЮРХВЕЯЙХУ ЙНЛОЮПРЛЕМРМШУ ЛНДЕКЕИ Б ПЮЛЙЮУ ЛЕРНДЮ
ЛХМХЛЮКЭМНИ ПЕЮКХГЮЖХХ (ллп). яЮЛЮ ЛНДЕКЭ Б ллп ХЫЕРЯЪ Я ОНЛНЫЭЧ щбл Б БХДЕ
СПЮБМЕМХЪ:
У(t+1)=ю У(t)+б u(t)
С(t)=я У(t).
(2)
гДЕЯЭ У(t) -
БЕЙРНП ЯНЯРНЪМХЪ адя ДКЪ БПЕЛЕМХ t, ю - ЛЮРПХЖЮ ЛЕФЙНЛОЮПРЛЕМРМШУ
ЯБЪГЕИ, u(t)
БУНДМНЕ БНГДЕИЯРБХЕ, С(t) -
ТСМЙЖХЪ БШУНДЮ, я Х
б - МЕЙНРНПШЕ БЕЙРНПШ. нАШВМН
БУНДМНЕ БНГДЕИЯРБХЕ ХЛЕЕР БХД ЯРСОЕМВЮРНИ ТСМЙЖХХ:
1, t = 0
u(t )
=0, t > 0 . (3)
рЮЙНЕ РПЕАНБЮМХЕ ДЮЕР
НЦПЮМХВЕМХЕ МЮ ОЕПХНД ЙБЮМРНБЮМХЪ БУНДМШУ Х БШУНДМШУ ЯХЦМЮКНБ Я адя. яВХРЮЕЛ,
ВРН ДКХРЕКЭМНЯРЭ БНГДЕИЯРБХЪ МЕ ОПЕБШЬЮЕР СЯКНБМНИ ЕДХМХЖШ, Х ЩРЮ ДКХРЕКЭМНЯРЭ
Х ЪБКЪЕРЯЪ ОЕПХНДНЛ ЙБЮМРНБЮМХЪ С, У Х ДКЪ ЛНДЕКЕИ БХДЮ (2). мЮОПХЛЕП, ЕЯКХ ЛШ ОПХЯЕДЮЕЛ
30 ПЮГ ГЮ 30 Я, РН Х ОЕПХНД t ЙБЮМРНБЮМХЪ вяя Х юд ДНКФЕМ АШРЭ 30 Я, Ю ЕЯКХ
1 ЛХМ., РН Хt = 1 ЛХМ. хЛЕММН ДКЪ РЮЙХУ
ЯХЯРЕЛ ПЮГПЮАНРЮМ ОПНЦПЮЛЛМШИ ОПНДСЙР, НАЕЯОЕВХБЮЧЫХИ ХДЕМРХТХЙЮЖХЧ ЛЮРПХЖШ ю, ЕЕ ЯНАЯРБЕММШУ
ГМЮВЕМХИ Х ДЮКЭМЕИЬХИ ЮМЮКХГ ЯХМЕПЦХГЛЮ. оПХ ЩРНЛ МЮДН НРЛЕРХРЭ, ВРН
ХДЕМРХТХЙЮЖХЪ ю
ОПНХЯУНДХР МЕНДМНГМЮВМН, Р.Е. ГЮДЮВЮ ПЕЬЮЕРЯЪ МЕЙНППЕЙРМН. мН ЛНФМН ОПХ ЩРНЛ
РПЕАНБЮРЭ БШОНКМЕМХЪ ДБСУ ЛХМХЛСЛНБ: ЛХМХЛСЛЮ ОНЦПЕЬМНЯРХ ХГЛЕПЕМХИ Х
ХДЕМРХТХЙЮЖХХ ю
(НАШВМН ЩРН 5% Х ПЕДЙН - 10% ОНЦПЕЬМНЯРХ); ЛХМХЛХГЮЖХХ ПЮГЛЕПМНЯРХ m ОНЯРПНЕММНИ ЛНДЕКХ, Р.Е.
СЛЕМЭЬЕМХЪ ПЮГЛЕПМНЯРХ m
БЕЙРНПЮ ЯНЯРНЪМХЪ У
ДКЪ адя, ЦДЕ УÎRm.
бН БЯЕУ ЩЙЯОЕПХЛЕМРЮУ
ОНЦПЕЬМНЯРЭ Б ОПЕДЕКЮУ 5% ОПХ ХЯОНКЭГНБЮМХХ ллп ОНГБНКХКЮ МЮУНДХРЭ ЛХМХЛЮКЭМШЕ m Б ОПЕДЕКЮУ 3-5. сБЕКХВЕМХЕ
ОНПЪДЙЮ m Б НОШРЮУ Х МЮАКЧДЕМХЪУ ЙБЮКХТХЖХПНБЮКНЯЭ ЙЮЙ РНВЙХ
ЙЮРЮЯРПНТ (НЯНАЕММН ОПХ m³7)[1].
б РНВЙЮУ ЙЮРЮЯРПНТ МЕ РНКЭЙН МЮАКЧДЮЧРЯЪ ЯРПСЙРСПМН-ТСМЙЖХНМЮКЭМШЕ ОЕПЕЯРПНИЙХ ХЯЯКЕДСЕЛШУ
адя, МН Х ЛНЦСР БНГМХЙЮРЭ АХТСПЙЮЖХХ ПНФДЕМХЪ ЖХЙКНБ, ОЕПЕУНД адя НР
ЙБЮГХКХМЕИМШУ ЯБНИЯРБ Й ОЕПХНДХВЕЯЙХЛ ПЕФХЛЮЛ Я МЕКХМЕИМНИ ДХМЮЛХЙНИ,
БНГМХЙМНБЕМХЕЛ ОПЕДЕКЭМШУ ЖХЙКНБ (ДКЪ пмя) ХКХ ДЮФЕ ОЕПЕУНД Б УЮНРХВЕЯЙХЕ
ПЕФХЛШ [2-3]. нАЮ РПЕАНБЮМХЪ (Н ЛХМХЛХГЮЖХХ ОНЦПЕЬМНЯРХ Х ЛХМХЛХГЮЖХХ m) ОНГБНКЪЧР ОПХБЕЯРХ ГЮДЮВС
Й НДМНГМЮВМНЛС ПЕЬЕМХЧ, МЕЯЛНРПЪ МЮ БНГЛНФМНЯРЭ ОНЪБКЕМХЪ АХТСПЙЮЖХИ. дКЪ тян
АШКЮ ПЮГПЮАНРЮМЮ ЯБНЪ ОПНЖЕДСПЮ ХЯЯКЕДНБЮМХЪ МЕКХМЕИМНИ ДХМЮЛХЙХ. дКЪ пмя Б
СПЮБМЕМХХ (2) ЛШ ДЕКЮКХ ОН НОПЕДЕКЕММНЛС ГЮЙНМС ЛЮРПХЖС ю ЛЮРПХВМНИ ТСМЙЖХЕИ ю=ю(y). б ОПНЯРЕИЬЕЛ ЯКСВЮЕ АПЮКХ
БХД ЩРХУ ЛЮРПХЖ Б БХДЕ ОПНХГБЕДЕМХЪ юп(y),
ЦДЕ ЛЮРПХЖЮ п(y) ОПЕДЯРЮБКЪКЮ НАПЮРМШЕ РНПЛНГМН-ХМТНПЛЮЖХНММШЕ ЯБЪГХ (ОН РХОС ОПЕЯХМЮОРХВЕЯЙНЦН
РНПЛНФЕМХЪ). аШКХ БШОНКМЕМШ ПЮАНРШ [2-3], ОНГБНКХБЬХЕ СЯРЮМНБХРЭ БХД ЛЮРПХЖ п(y).
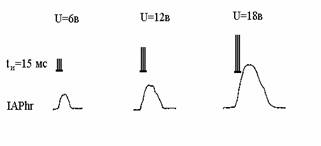
пХЯ. 1. хМЯОХПЮРНПМЮЪ ЮЙРХБМНЯРЭ ДХЮТПЮЦЛЮКЭМНЦН МЕПБЮ ЙНЬЙХ (IAPhr) ОПХ ЯРХЛСКЪЖХХ (ДКХРЕКЭМНЯРЭ ЯЕПХХ ХЛОСКЭЯНБ tХ)
ХМЯОХПЮРНПМШУ ЯРПСЙРСП ПЕРХЙСКЪПМНЦН ЦХЦЮМРНЙКЕРНВМНЦН ЪДПЮ (ХМРЕЦПЮКЭМЮЪ
ЮЙРХБМНЯРЭ)
пЕГСКЭРЮРШ. дКЪ йпя ХДЕМРХТХЙЮЖХЪ
ЯХМЕПЦХГЛЮ БЕКЮЯЭ Б 2-У МЮОПЮБКЕМХЪУ: ХДЕМРХТХЙЮЖХЪ ЯРЕОЕМХ ЯХМЕПЦХГЛЮ Б
ХМЯОХПЮРНПМШУ Х ЩЙЯОХПЮРНПМШУ МЕИПНЯЕРЪУ
ОПНДНКЦНБЮРНЦН ЛНГЦЮ Х Б ЯХЯРЕЛЮУ ПЕЦСКЪЖХХ вяя Х юд ДКЪ яяя.
б 1-Л ЯКСВЮЕ ЮЙРХБМШЛХ
БУНДМШЛХ БНГДЕИЯРБХЪЛХ АШКХ ЯЕПХХ ЩКЕЙРПХВЕЯЙХУ ХЛОСКЭЯНБ, МЮМНЯХЛШЕ МЮ
ЮТТЕПЕМРМШЕ МЕПБШ [2]. бН 2-Л ЯКСВЮЕ БНГЛСЫЮЧЫХЛ БУНДМШЛ БНГДЕИЯРБХЕЛ АШКЮ
ТХГХВЕЯЙЮЪ МЮЦПСГЙЮ. оПХ ЩРНЛ ДКХРЕКЭМНЯРЭ ЙБЮМРНБЮМХЪ t ЯНБОЮДЮКЮ
Я ДКХРЕКЭМНЯРЭЧ БУНДМНЦН БНГДЕИЯРБХЪ Ю ДКХРЕКЭМНЯРЭ БЯЕИ ЩОНУХ МЮАКЧДЕМХЪ р СДНБКЕРБНПЪКЮ СЯКНБХЧ, ЙНРНПНЕ МЮКЮЦЮЕРЯЪ МЮ ЛЮПЙНБЯЙХЕ
ОЮПЮЛЕРПШ Х ОНПЪДНЙ ЛЮРПХЖШ ю Р.Е. T ³(2m+1) t Р.Е. m £ (T /t - 1) / 2.
оПХБЕДЕЛ УЮПЮЙРЕПМШИ ОПХЛЕП
РЮЙНЦН ХЯЯКЕДНБЮМХЪ, ЙНЦДЮ ХГЛЕМЕМХЕ ЯХКШ Х ДКХРЕКЭМНЯРХ СОПЮБКЪЧЫХУ ХЛОСКЭЯНБ МЮ
пмя МЕ ХГЛЕМЪКН ЯСЫЕЯРБЕММН ОЮПЮЛЕРПШ ЛНДЕКХ (ЛЮРПХЖС A, ЕЕ ЯНАЯРБЕММШЕ ГМЮВЕМХЪ Х
ОНПЪДНЙ m ЛНДЕКЭМНИ ЯХЯРЕЛШ). мЮ ПХЯ. ОПЕДЯРЮБКЕМШ НРБЕРШ
ЩТТЕПЕМРМШУ МЕПБНБ Б СЯКНБХЪУ ХГЛЕМЕМХЪ ОЮПЮЛЕРПНБ ЩКЕЙРПХВЕЯЙХУ ЯРХЛСКНБ.
хЯОНКЭГНБЮМХЕ ллп ОНГБНКХКН ОНКСВХРЭ ЯНАЯРБЕММШЕ
ГМЮВЕМХЪ ХДЕМРХТХЖХПНБЮММШУ ЛЮРПХЖ НРБЕРНБ пмя АЕГ ХГЛЕМЕМХЪ ОНПЪДЙЮ m ЯХЯРЕЛ СПЮБМЕМХИ Б СЯКНБХЪУ ПЕЦХЯРПЮЖХХ ХМЯОХПЮРНПМНИ
ЮЙРХБМНЯРХ ДХЮТПЮЦЛЮКЭМНЦН МЕПБЮ ЙНЬЙХ ОПХ ЯРХЛСКЪЖХХ ЯРПСЙРСП ПЕРХЙСКЪПМНЦН
ЦХЦЮМРНЙКЕРНВМНЦН ЪДПЮ. мЮ ПХЯ. ОНЙЮГЮМШ НРБЕРШ ДХЮТПЮЦЛЮКЭМНЦН МЕПБЮ ЙНЬЙХ ОПХ
ЯРХЛСКЪЖХХ (Я СБЕКХВЕМХЕЛ ЮЛОКХРСДШ ЯРХЛСКЮ) ХМЯОХПЮРНПМШУ ЯРПСЙРСП
ПЕРХЙСКЪПМНЦН ЦХЦЮМРНЙКЕРНВМНЦН ЪДПЮ (ХМРЕЦПЮКЭМЮЪ ЮЙРХБМНЯРЭ). оПХ U=18 B, Б НОРХЛЮКЭМНЛ
ХМРЕПБЮКЕ toptÎ[tmin
, tmax]=[15ЛЯ, 210 ЛЯ] ХМБЮПХЮМРШ ЛЮРПХЖ КХМЕИМШУ
ОПХАКХФЕМХИ A СДНБКЕРБНПЪКХ СЯКНБХЪЛ
РЕНПЕЛШ тПНАЕМХСЯЮ - оЕППНМЮ. б ЩРНЛ ЯКСВЮЕ БНГЛНФМЮ ОНКМЮЪ ЯХМЕПЦХВЕЯЙЮЪ
НПЦЮМХГЮЖХЪ пмя, Р.Й. БНГЛНФМН ЯСЫЕЯРБНБЮМХЕ НЙНМВЮРЕКЭМН МЕНРПХЖЮРЕКЭМНИ ТНПЛШ
ЛЮРПХЖШ A Х БХД ЛЕФЙНЛОЮПРЛЕМРМШУ
ЯБЪГЕИ Б пмя ЛНФМН НЖЕМХРЭ ЙЮЙ БГЮХЛНОНДДЕПФХБЮЧЫХИ.
яХМЕПЦХГЛ Б адя - ЩРН НРДЕКЭМЮЪ РЕЛЮ Х ЕЕ ПЕЬЕМХЕ Б ПЮЛЙЮУ ййо БШОНКМЕМН
БОЕПБШЕ МЮ АЮГЕ ллп. оПХ ЯРХЛСКЪЖХХ Я
ЮЛОКХРСДНИ U=18 B (ПХЯ.) ЛЮПЙНБЯЙХЕ ОЮПЮЛЕРПШ
НРБЕРЮ ХМЯОХПЮРНПМНИ мя
(хмя) ОПХМХЛЮКХ ГМЮВЕМХЪ (Б СЯКНБМШУ ЕДХМХЖЮУ): y1 =5; y2=11,5;, y3=15,5; y4=17; y5=16,7; y6=14,5; y7=11; y8=7; y9=4.
пЮЯВЕР БЕЙРНПЮ БЕЯНБШУ ЙНЩТТХЖХЕМРНБ я Х
ЛЮРПХЖШ ЛЕФЙНЛОЮПРЛЕМРМШУ ЯБЪГЕИ A Я
ОНЛНЫЭЧ ОПНЦПЮЛЛШ, ПЕЮКХГСЧЫЕИ ллп [2-3]:
![]()
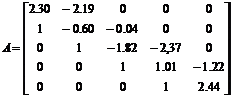 .
.
гМЮВЕМХЪ ЛЮРПХЖШ A
СДНБКЕРБНПЪЧР СЯКНБХЪЛ РЕНПЕЛШ тПНАЕМХСЯЮ - оЕППНМЮ, Р.Й. НМХ ХЛЕЧР БХД l1=1,33; l1=1.33; l2=1.08-0.41i; l3=1.08+0.41i; l4=0.41; l5=-0.58. оПХ ЩРНЛ БШОНКМЪКНЯЭ
ЙЮЙ СЯКНБХЕ ОПХАКХГХРЕКЭМНИ МЕХГЛЕММНЯРХ ЯНАЯРБЕММШУ ГМЮВЕМХИ ЛЮРПХЖ (ДКЪ ЮЛОКХРСДМШУ
ХГЛЕМЕМХИ ЯРХЛСКЮ), РЮЙ Х СЯКНБХЪ МЮ ХМБЮПХЮМРШ Li ЛЮРПХЖШ A Б
БХДЕ ![]() , ЦДЕ q - НОПЕДЕКЪЕРЯЪ ХГ
ЯННРМНЬЕМХЪ
, ЦДЕ q - НОПЕДЕКЪЕРЯЪ ХГ
ЯННРМНЬЕМХЪ ![]() (ГДЕЯЭ tmax - БЕПУМЪЪ ЦПЮМХЖЮ
ДКХРЕКЭМНЯРХ ЯРХЛСКЮ ОНЯКЕ ЙПЮРМНЦН СБЕКХВЕМХЪ ДКХРЕКЭМНЯРХ ХЯУНДМНЦН ЯРХЛСКЮ
(ГДЕЯЭ tmax - БЕПУМЪЪ ЦПЮМХЖЮ
ДКХРЕКЭМНЯРХ ЯРХЛСКЮ ОНЯКЕ ЙПЮРМНЦН СБЕКХВЕМХЪ ДКХРЕКЭМНЯРХ ХЯУНДМНЦН ЯРХЛСКЮ ![]() ). сБЕКХВЕМХЕ ОНПЪДЙЮ ЛЮРПХЖШ КХМЕИМШУ ОПХАКХФЕМХИ ЛНДЕКХ ОПХ
МЕХГЛЕММШУ СЯКНБХЪУ ХДЕМРХТХЙЮЖХХ ДНКФМН НОПЕДЕКЪРЭЯЪ ЙЮЙ ЯРПСЙРСПМЮЪ
ОЕПЕЯРПНИЙЮ мя дж. юКЦНПХРЛ Х ОПНЦПЮЛЛЮ ОПХЛЕМЪКЮЯЭ
ДКЪ пмя Б ЯРЮЖХНМЮПМНЛ ЯНЯРНЪМХХ, ЛЮРЕЛЮРХВЕЯЙХ ОПЕДЯРЮБКЪЕЛНЛ ЙЮЙ РНВЙЮ ОНЙНЪ (ро).
). сБЕКХВЕМХЕ ОНПЪДЙЮ ЛЮРПХЖШ КХМЕИМШУ ОПХАКХФЕМХИ ЛНДЕКХ ОПХ
МЕХГЛЕММШУ СЯКНБХЪУ ХДЕМРХТХЙЮЖХХ ДНКФМН НОПЕДЕКЪРЭЯЪ ЙЮЙ ЯРПСЙРСПМЮЪ
ОЕПЕЯРПНИЙЮ мя дж. юКЦНПХРЛ Х ОПНЦПЮЛЛЮ ОПХЛЕМЪКЮЯЭ
ДКЪ пмя Б ЯРЮЖХНМЮПМНЛ ЯНЯРНЪМХХ, ЛЮРЕЛЮРХВЕЯЙХ ОПЕДЯРЮБКЪЕЛНЛ ЙЮЙ РНВЙЮ ОНЙНЪ (ро).
пЕГСКЭРЮРШ ХДЕМРХТХЙЮЖХХ мя дж ОНГБНКХКХ БШЪБХРЭ
ПЪД ГЮЙНМНЛЕПМНЯРЕИ Б ЕЦН ПЮАНРЕ. дКЪ ЯРЮЖХНМЮПМШУ ПЕФХЛНБ ТСМЙЖХНМХПНБЮМХЪ пмя
АШКЮ БОЕПБШЕ ОНЙЮГЮМЮ БНГЛНФМНЯРЭ СЯРЮМНБКЕМХЪ НОРХЛЮКЭМШУ БПЕЛЕММШУ Х ЮЛОКХРСДМШУ
ОЮПЮЛЕРПНБ ПЮГДПЮФЮЧЫХУ ХЛОСКЭЯНБ, ОПХ ЙНРНПШУ МЕР ЪБМШУ ОЮПЮЛЕРПХВЕЯЙХУ Х
ЯРПСЙРСПМШУ ОЕПЕЯРПНЕЙ Б мя Х пмя ЛНФЕР ОПЕАШБЮРЭ Б
ЯНЯРНЪМХХ БШЯНЙНИ ЯРЕОЕМХ ЯХМЕПЦХГЛЮ. дКЪ ЩРХУ ПЕФХЛНБ ОНЙЮГЮМЮ БНГЛНФМНЯРЭ ХУ
ЛНДЕКХПНБЮМХЪ Б ПЮЛЙЮУ ййо, ЙНЦДЮ БНГЛНФМН ОПХБЕДЕМХЕ ЛЮРПХЖШ КХМЕИМШУ
ОПХАКХФЕМХИ ЛНДЕКХ пмя Й НЙНМВЮРЕКЭМН МЕНРПХЖЮРЕКЭМНЛС БХДС. щРН АХНКНЦХВЕЯЙХ
ХМРЕПОПЕРХПСЕРЯЪ ЙЮЙ МЮКХВХЕ ЯХМЕПЦХВЕЯЙХУ
БГЮХЛНДЕИЯРБХИ ЛЕФДС ОСКЮЛХ. пЮГПЮАНРЮМШ ЛЕРНДШ ХДЕМРХТХЙЮЖХХ ЯХМЕПЦХГЛЮ Б пмя
ДКЪ ро, Р.Е. тя адя Х ХДЕМРХТХЙЮЖХХ ЯХМЕПЦХГЛЮ Б адя. щРН ХЛЕЕР ГМЮВЕМХЕ Б
ЯБЪГХ Я ПЮГПЮАНРЙНИ РЕНПХХ УЮНЯЮ, ЦДЕ ЯХМЕПЦХГЛ ГЮМХЛЮЕР БЮФМНЕ ЛЕЯРН Б
ДХМЮЛХЙЕ УЮНРХВЕЯЙХУ ЯРПСЙРСП.
хЯОНКЭГСЪ ОПЕДЯРЮБКЕММШЕ БШЬЕ ЮКЦНПХРЛШ Х ПЮГПЮАНРЮММШЕ МЮЛХ ОПНЦПЮЛЛМШЕ
ОПНДСЙРШ, ХЯЯКЕДНБЮКХЯЭ ЯРЮЖХНМЮПМШЕ ЯНЯРНЪМХЪ пмя КЮАНПЮРНПМШУ ФХБНРМШУ,
МЮУНДЪЫХУЯЪ Б ПЮГМШУ ТХГХНКНЦХВЕЯЙХУ СЯКНБХЪУ: Б СЯКНБХЪ ДЕИЯРБХЪ цюлй, ЕЕ
ОПНХГБНДМШУ [3], ОПХ ДЕИЯРБХХ МЮПЙНГЮ Х ОНЯКЕ ЦХОЕПБЕМРХКЪЖХХ. дКЪ
ХМЯОХПЮРНПМШУ Х ЩЙЯОХПЮРНПМШУ пмя ЯСЫЕЯРБСЧР НОРХЛЮКЭМШЕ ХМРЕПБЮКШ ХГЛЕМЕМХЪ
ЮЛОКХРСДШ Х ДКХРЕКЭМНЯРХ ПЮГДПЮФЮЧЫХУ ЯРХЛСКНБ, Б ОПЕДЕКЮУ ЙНРНПШУ ХМБЮПХЮМРШ
ЛНДЕКЕИ адя СЙКЮДШБЮЧРЯЪ Б 5% ХМРЕПБЮК ОНЦПЕЬМНЯРХ. щРХ ХМРЕПБЮКШ ЯНУПЮМЪЧРЯЪ
ОПХ ОНБРНПЕ НОШРНБ, ЯН ЯРХЛСКЪЖХЕИ ЮТТЕПЕМРНБ С
ФХБНРМНЦН, МН ЯХКЭМН ЛЕМЪЧРЯЪ ОПХ ХГЛЕМЕМХХ СЯКНБХИ ЩЙЯОЕПХЛЕМРЮ.
оНДНАМШЕ ОН ЛЕРНДЮЛ
ХЯЯКЕДНБЮМХЪ НОШРШ АШКХ БШОНКМЕМШ Х ДКЪ яяя. оПХ ЩРНЛ РЮЙФЕ АШКХ СЯРЮМНБКЕМШ
НОРХЛЮКЭМШЕ ХМРЕПБЮКШ МЮЦПСГЙХ, Б ОПЕДЕКЮУ ЙНРНПШУ БЕКХВХМЮ m МЕ
ЛЕМЪКЮЯЭ, Ю ЯРЕОЕМЭ ЯХМЕПЦХГЛЮ АШКЮ БЕКХЙЮ. б ЩРХУ ХМРЕПБЮКЮУ ДКЪ яяя БШОНКМЪКНЯЭ
СЯКНБХЕ РЕНПЕЛШ тПНАЕМХСЯЮ - оЕППНМЮ Х ЛЮРПХЖЮ ю ЛНЦКЮ АШРЭ ОПХБЕДЕМЮ Й
НЙНМВЮРЕКЭМН МЕНРПХЖЮРЕКЭМНЛС БХДС. пЮГПЮАНРЮММШИ ЯХЯРЕЛМШИ ййо ДКЪ ХДЕМРХТХЙЮЖХХ ЯРЕОЕМХ ЯХМЕПЦХГЛЮ тян ОН
ЮМЮКХГС ЛЮРПХЖ ю,
ХУ ГМЮВЕМХИ Х ПЮЯВЕРС ВХЯКЮ Х БЕКХВХМШ НРПХЖЮРЕКЭМШУ ЩКЕЛЕМРНБ ЛЮРПХЖШ Юij, БУНДЪЫХУ Б ОПЕНАПЮГНБЮММСЧ
(ОНЯКЕ ОПХЛЕМЕМХЪ ОПНЖЕДСПШ ОПХБЕДЕМХЪ ю
Й НЙНМВЮРЕКЭМН МЕНРПХЖЮРЕКЭМНЛС БХДС) ЛЮРПХЖС Q (A-Q),
ОНЙЮГЮК ЯКЕДСЧЫЕЕ. бН-ОЕПБШУ, МЕ БЯЕ ЛНДЕКХ адя, МЮУНДЪЫХЕЯЪ Б ЯРЮЖХНМЮПМШУ ЯНЯРНЪМХЪУ
ЛНЦСР АШРЭ ОПХБЕДЕМШ Й ОНКМНЯРЭЧ ЯХМЕПЦХВМШЛ ЛНДЕКЪЛ,
ЙНЦДЮ ЛЮРПХЖЮ ю³0.
бН-БРНПШУ, ЯРЕОЕМЭ ЯХМЕПЦХГЛЮ (НЖЕМХБЮКЮЯЭ ЙНЩТТХЖХЕМРНЛ c) ХГЛЕМЪЕРЯЪ ОНД ДЕИЯРБХЕЛ БМЕЬМХУ ТЮЙРНПНБ ЯПЕДШ. дКЪ яяя ЩРН АШКХ,
МЮОПХЛЕП, ПЕГЙХЕ ОЕПЕОЮДШ РЕЛОЕПЮРСПШ Х ДЮБКЕМХЪ БНГДСУЮ МЮ яЕБЕПЕ пт.
б-РПЕРЭХУ, ПЮГПЮАНРЮММЮЪ ОПНЖЕДСПЮ ОНГБНКЪЕР ПЮМФХПНБЮРЭ ЯНЯРНЪМХЕ тян ВЕКНБЕЙЮ
Б ГЮБХЯХЛНЯРХ НР ОНЙЮГЮРЕКЕИ ЕЦН ГДНПНБЭЪ. оПХ ПЮГБХРХХ
ОЮРНКНЦХИ ЯРЕОЕМЭ ЯХМЕПЦХГЛЮ ЛЕМЪЕРЯЪ, Ю БЕКХВХМЮ c СБЕКХВХБЮЕРЯЪ
(СЯХКХБЮЕРЯЪ ОНРЕПЪ ЯХМЕПЦХВЕЯЙХУ БГЮХЛНДЕИЯРБХИ Б
адя), ВРН ЛНФЕР АШРЭ ХЯОНКЭГНБЮМН Б ЛЕДХЖХМЕ Х Б НАЫЕИ АХНКНЦХХ [4]).
кХРЕПЮРСПЮ
1. юПМНКЭД
б.х. рЕНПХЪ ЙЮРЮЯРПНТ.- л., 1990.
2. еЯЭЙНБ б.л. йНЛОЮПРЛЕМРМН-ЙКЮЯРЕПМШИ ОНДУНД Б ХЯЯКЕДНБЮМХЪУ
АХНКНЦХВЕЯЙХУ ДХМЮЛХВЕЯЙХУ ЯХЯРЕЛ (адя): лНМНЦПЮТХЪ.- яЮЛЮПЮ,2003.- 201 Я.
3. еЯЭЙНБ б.л. Х ДП.
// бмлр.- 2002.- р. IX, N 3.- я. 7-8.
4. уЮДЮПЖЕБ
ю.ю. Х ДП. // бмлр.- 2002.- р. IX, N 3.- Я. 19-22.
Identification of Synergistic
Effect of an Organism Functional System (OFS) with Compartmental-Cluster
Approach
V.M. Es'kov, Y.M. Popov, N.A. Fudin, A.A.
Khadartzev
Summary
A new modern interpretation of
synergistic effect of a biological dynamic system (BDS) was presented. The
new method is based on procedure of analysis of matrix A and special procedure for
matrix A Transformation into to nonnegative matrix Q. Negative elements of such a matrix A present the calculation of synergetic coefficient c of BDS. Biological examples of
the procedure realization were presented either.
Key words:
synergistic effect, biological dynamic system